

Alternative intermolecular contacts underlie the rotavirus VP5* two- to three-fold rearrangement
- PMID: 16511559
- PMCID: PMC1440311
- DOI: 10.1038/sj.emboj.7601034
Alternative intermolecular contacts underlie the rotavirus VP5* two- to three-fold rearrangement
Abstract
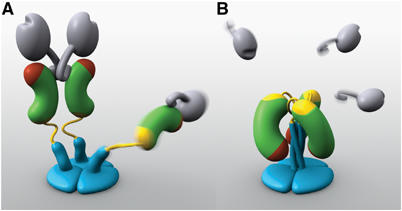
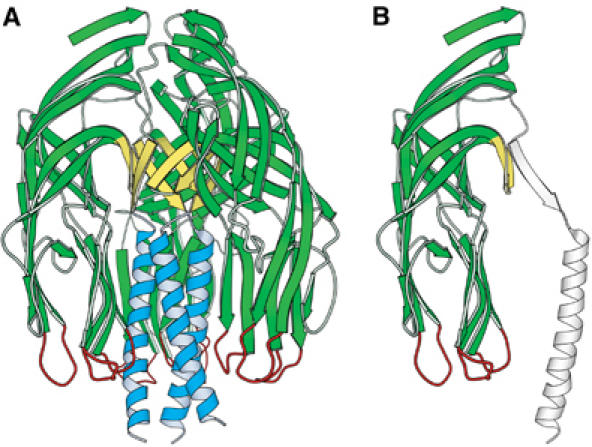
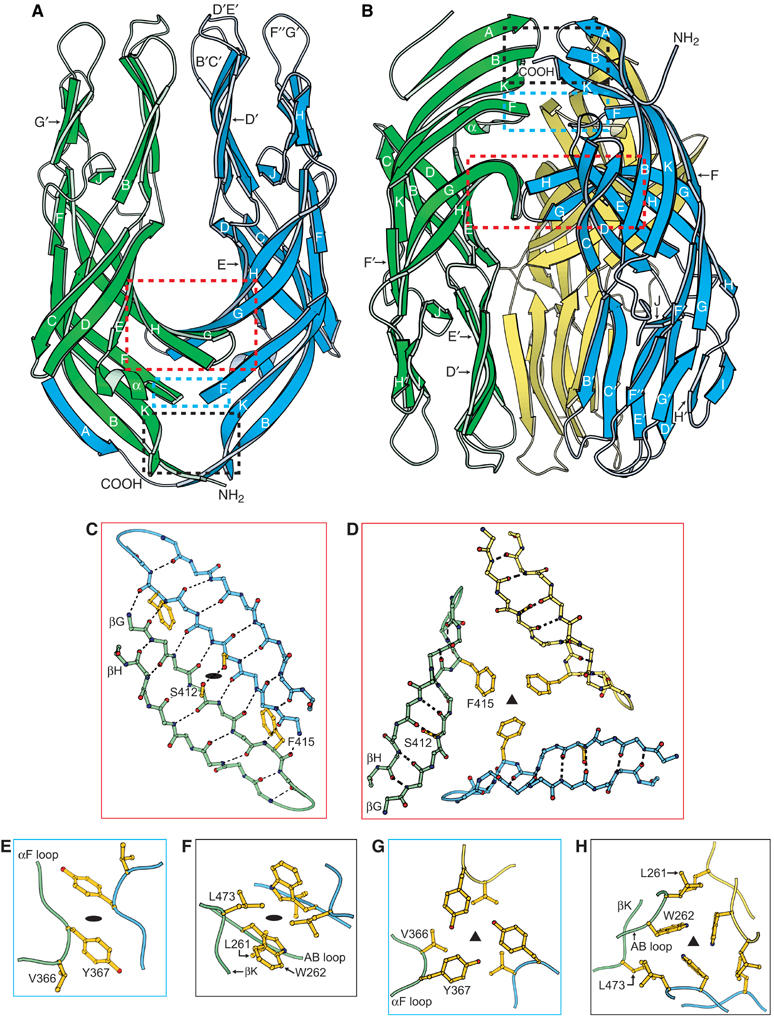
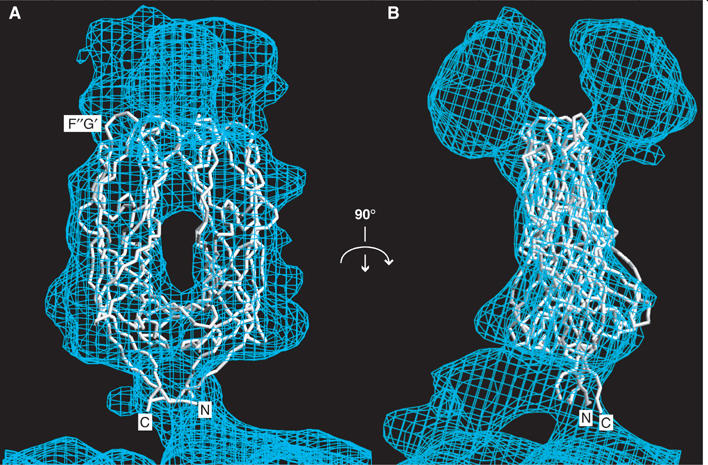
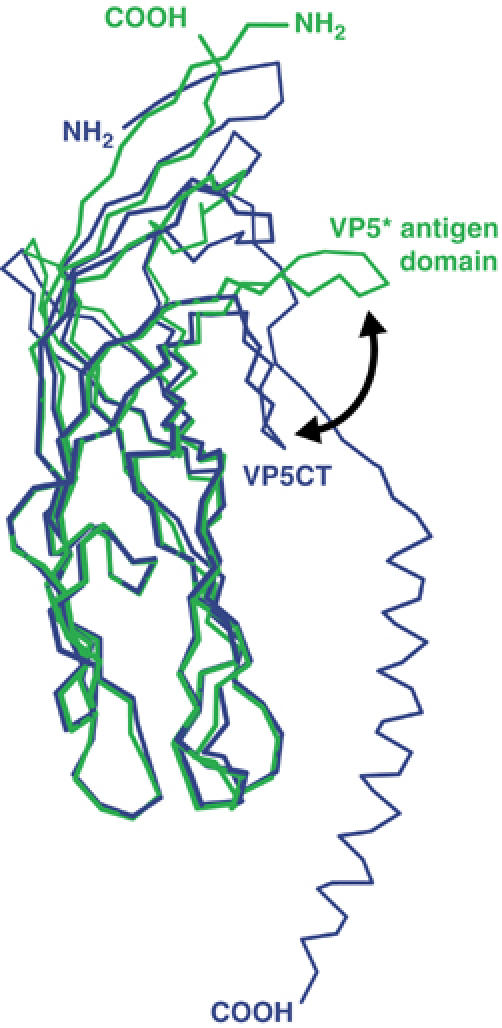
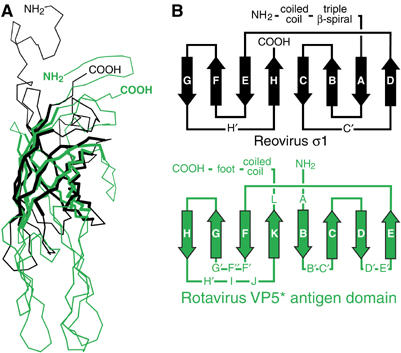
The spike protein VP4 is a key component of the membrane penetration apparatus of rotavirus, a nonenveloped virus that causes childhood gastroenteritis. Trypsin cleavage of VP4 produces a fragment, VP5*, with a potential membrane interaction region, and primes rotavirus for cell entry. During entry, the part of VP5* that protrudes from the virus folds back on itself and reorganizes from a local dimer to a trimer. Here, we report that a globular domain of VP5*, the VP5* antigen domain, is an autonomously folding unit that alternatively forms well-ordered dimers and trimers. Because the domain contains heterotypic neutralizing epitopes and is soluble when expressed directly, it is a promising potential subunit vaccine component. X-ray crystal structures show that the dimer resembles the spike body on trypsin-primed virions, and the trimer resembles the folded-back form of the spike. The same structural elements pack differently to form key intermolecular contacts in both oligomers. The intrinsic molecular property of alternatively forming dimers and trimers facilitates the VP5* reorganization, which is thought to mediate membrane penetration during cell entry.
Figures
References
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D 54: 905–921 - PubMed
-
- Collaborative Computational Project (1994) The CCP4 Suite: programs for protein crystallography. Acta Crystallogr D 50: 760–763 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
