

Drosophila Sex-lethal protein mediates polyadenylation switching in the female germline
- PMID: 16511567
- PMCID: PMC1422161
- DOI: 10.1038/sj.emboj.7601022
Drosophila Sex-lethal protein mediates polyadenylation switching in the female germline
Abstract
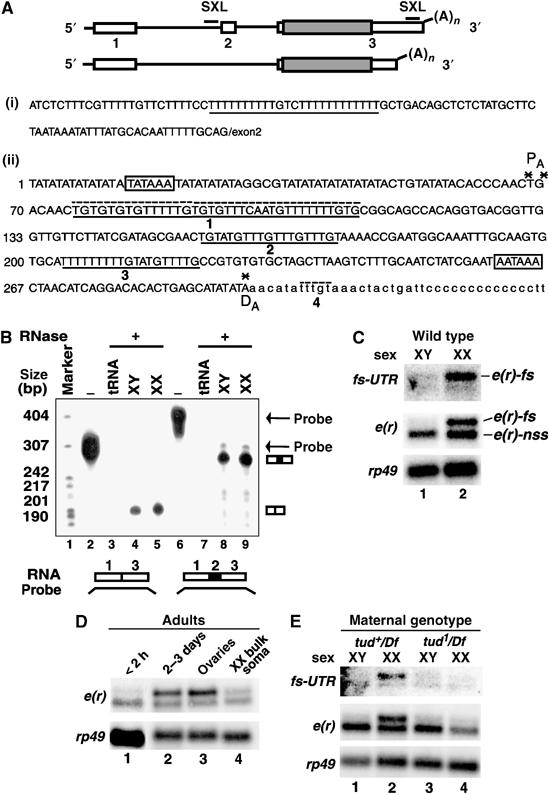
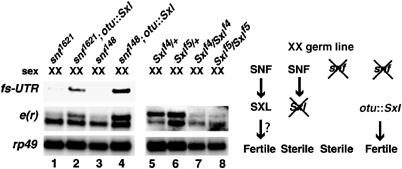
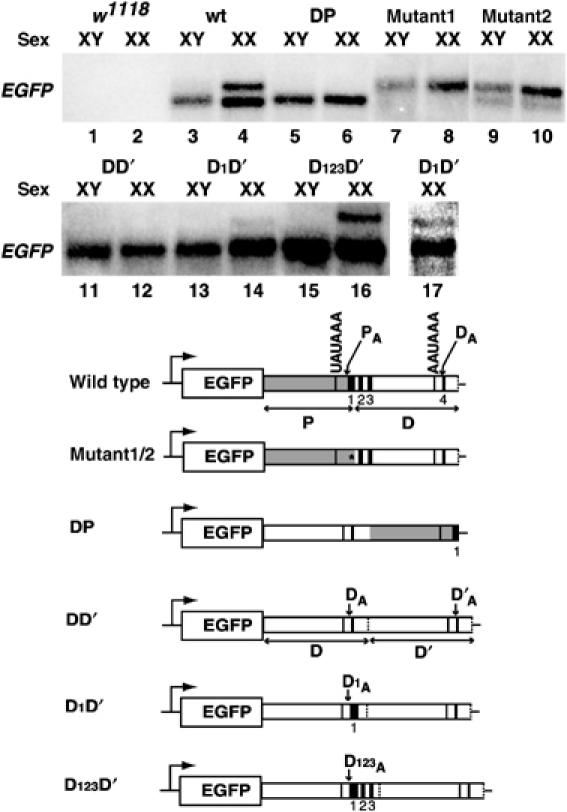
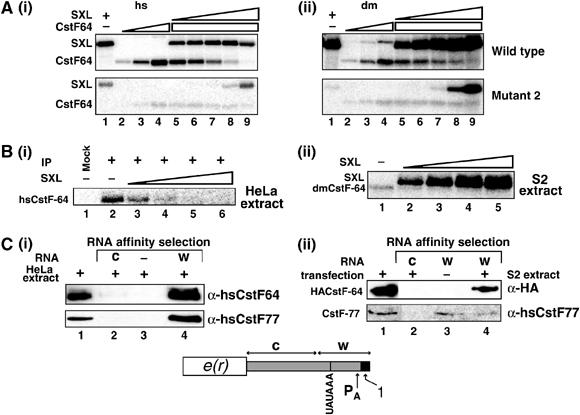
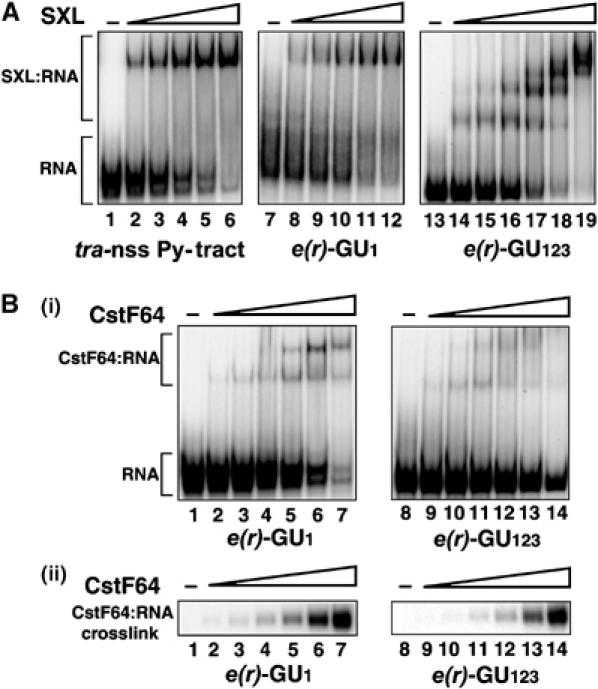
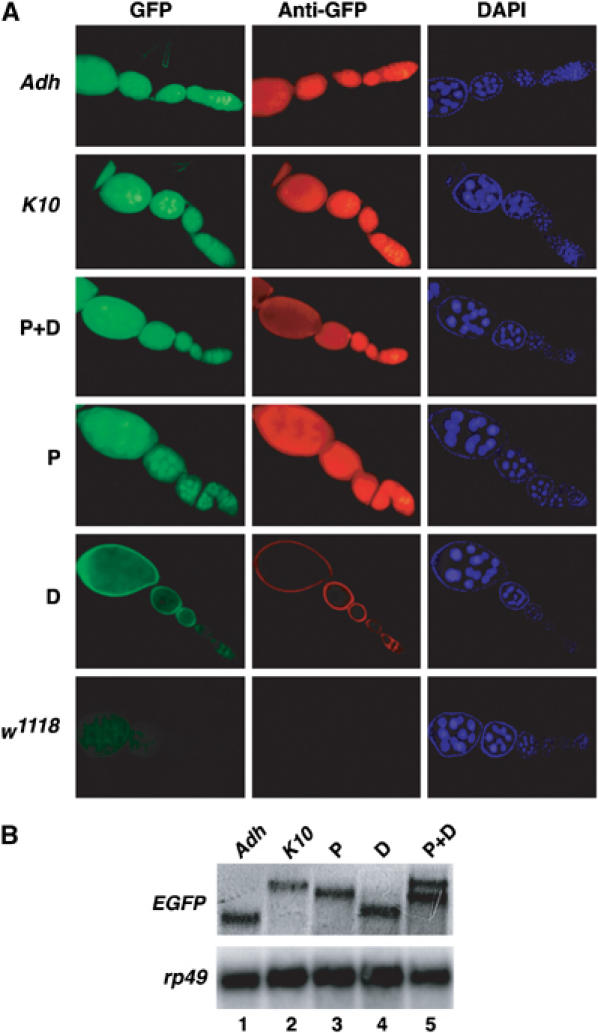
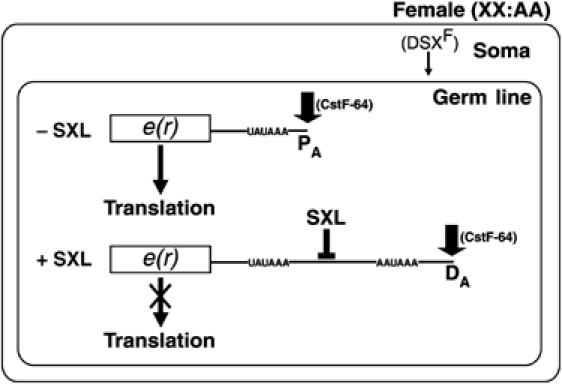
The Drosophila master sex-switch protein Sex-lethal (SXL) regulates the splicing and/or translation of three known targets to mediate somatic sexual differentiation. Genetic studies suggest that additional target(s) of SXL exist, particularly in the female germline. Surprisingly, our detailed molecular characterization of a new potential target of SXL, enhancer of rudimentary (e(r)), reveals that SXL regulates e(r) by a novel mechanism--polyadenylation switching--specifically in the female germline. SXL binds to multiple SXL-binding sites, which include the GU-rich poly(A) enhancer, and competes for the binding of CstF64 in vitro. The SXL-binding sites are able to confer sex-specific poly(A) switching onto an otherwise nonresponsive polyadenylation signal in vivo. The sex-specific poly(A) switching of e(r) provides a means for translational regulation in germ cells. We present a model for the SXL-dependent poly(A) site choice in the female germline.
Figures
References
-
- Amente S, Napolitano G, Licciardo P, Monti M, Pucci P, Lania L, Majello B (2005) Identification of proteins interacting with the RNAPII FCP1 phosphatase: FCP1 forms a complex with arginine methyltransferase PRMT5 and it is a substrate for PRMT5-mediated methylation. FEBS Lett 579: 683–689 - PubMed
-
- Black DL (2003) Mechanisms of alternative pre-messenger RNA splicing. Annu Rev Biochem 72: 291–336 - PubMed
-
- Bopp D, Bell LR, Cline TW, Schedl P (1991) Developmental distribution of female-specific Sex-lethal proteins in Drosophila melanogaster. Genes Dev 5: 403–415 - PubMed
-
- Bopp D, Horabin JI, Lersch RA, Cline TW, Schedl P (1993) Expression of the Sex-lethal gene is controlled at multiple levels during Drosophila oogenesis. Development 118: 797–812 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
