

Inefficient cytotoxic T lymphocyte-mediated killing of HIV-1-infected cells in vivo
- PMID: 16515366
- PMCID: PMC1395353
- DOI: 10.1371/journal.pbio.0040090
Inefficient cytotoxic T lymphocyte-mediated killing of HIV-1-infected cells in vivo
Abstract
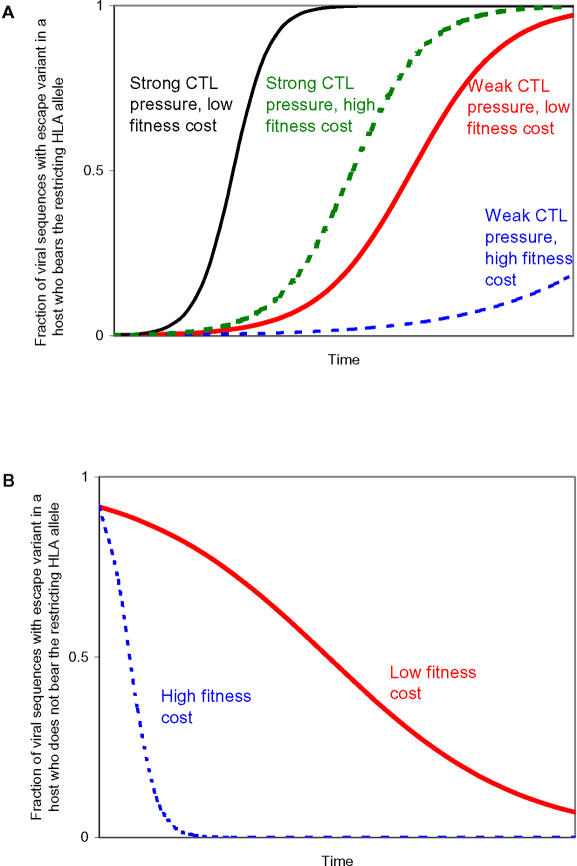
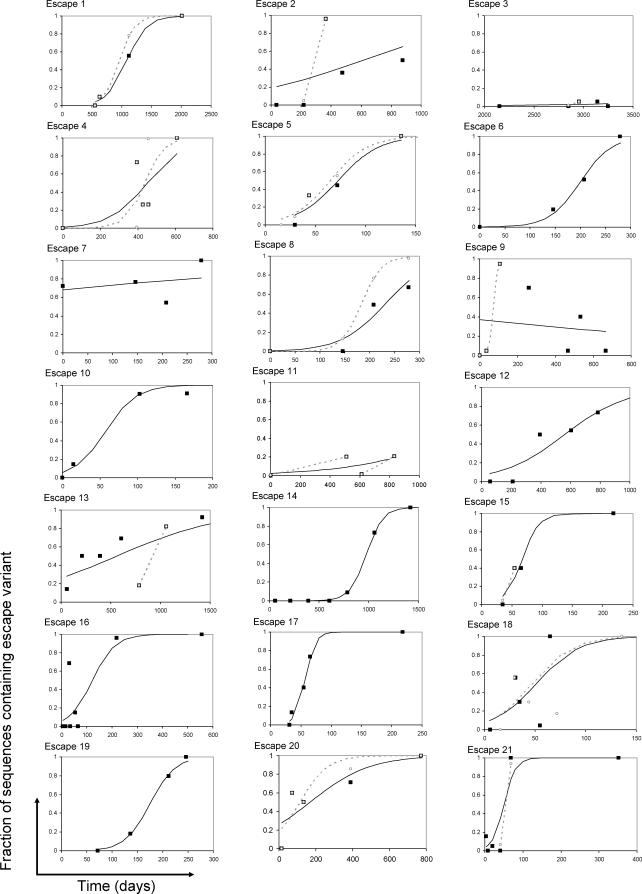
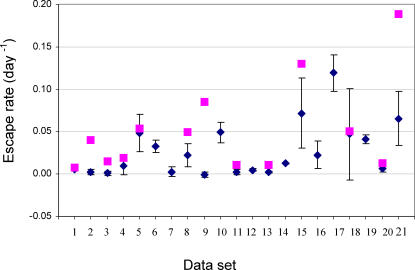
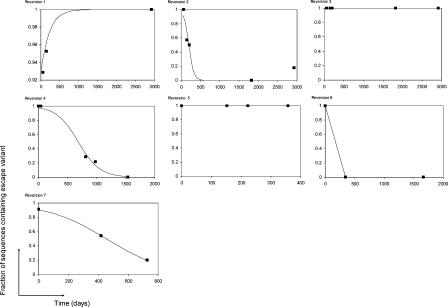
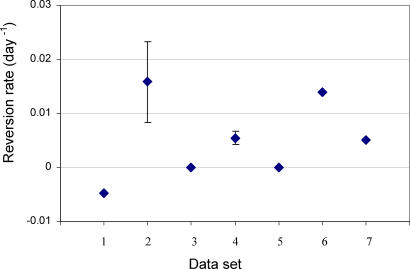
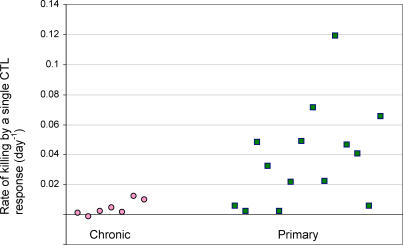
Understanding the role of cytotoxic T lymphocytes (CTLs) in controlling HIV-1 infection is vital for vaccine design. However, it is difficult to assess the importance of CTLs in natural infection. Different human leukocyte antigen (HLA) class I alleles are associated with different rates of progression to AIDS, indicating that CTLs play a protective role. Yet virus clearance rates following antiretroviral therapy are not impaired in individuals with advanced HIV disease, suggesting that weakening of the CTL response is not the major underlying cause of disease progression and that CTLs do not have an important protective role. Here we reconcile these apparently conflicting studies. We estimate the selection pressure exerted by CTL responses that drive the emergence of immune escape variants, thereby directly quantifying the efficiency of HIV-1-specific CTLs in vivo. We estimate that only 2% of productively infected CD4+ cell death is attributable to CTLs recognising a single epitope. We suggest that CTLs kill a large number of infected cells (about 10(7)) per day but are not responsible for the majority of infected cell death.
Figures
Comment in
-
Inefficient immune killer cells abet HIV infection.PLoS Biol. 2006 Apr;4(4):e114. doi: 10.1371/journal.pbio.0040114. Epub 2006 Mar 14. PLoS Biol. 2006. PMID: 20076553 Free PMC article. No abstract available.
References
-
- Wei X, Ghosh SK, Taylor ME, Johnson VA, Emini EA, et al. Viral dynamics in human immunodeficiency virus type 1 infection. Nature. 1995;373:117–122. - PubMed
-
- Ho DD, Neumann AU, Perelson AS, Chen W, Leonard JM, et al. Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature. 1995;373:123–126. - PubMed
-
- Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1 dynamics in vivo: Virion clearance rate, infected cell life-span, and viral generation time. Science. 1996;271:1582–1586. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
