

Reactive oxygen species play a role in regulating a fungus-perennial ryegrass mutualistic interaction
- PMID: 16517760
- PMCID: PMC1425850
- DOI: 10.1105/tpc.105.039263
Reactive oxygen species play a role in regulating a fungus-perennial ryegrass mutualistic interaction
Abstract
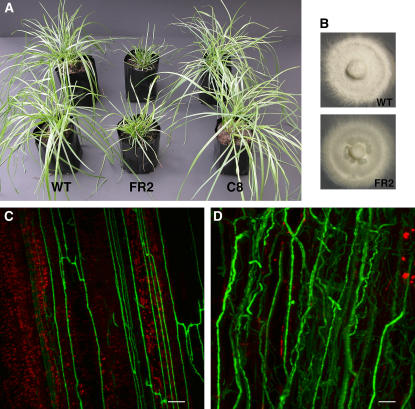
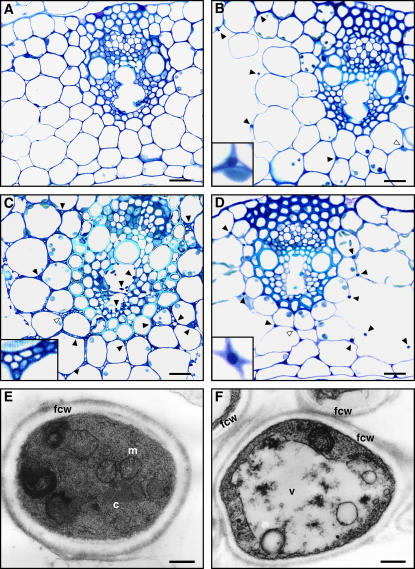
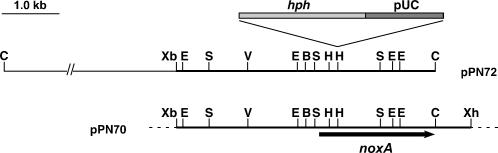
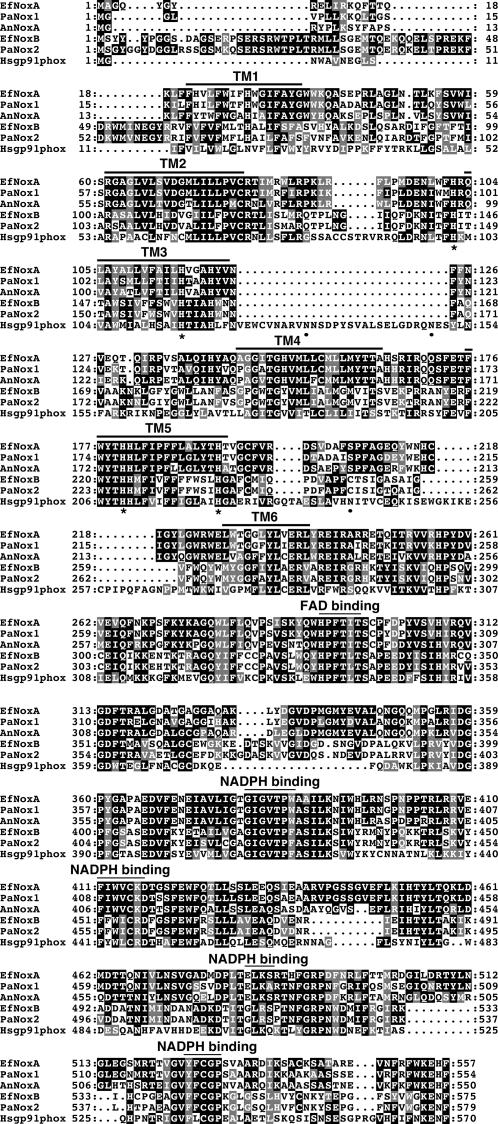
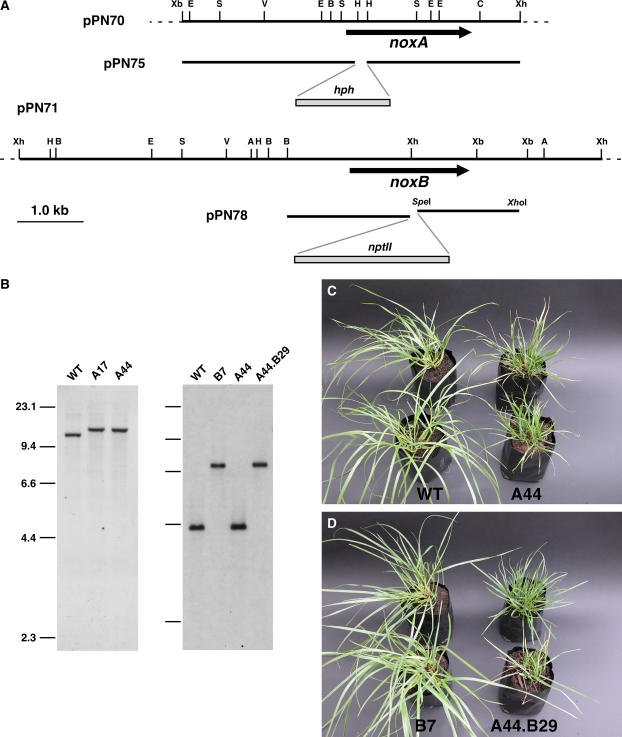
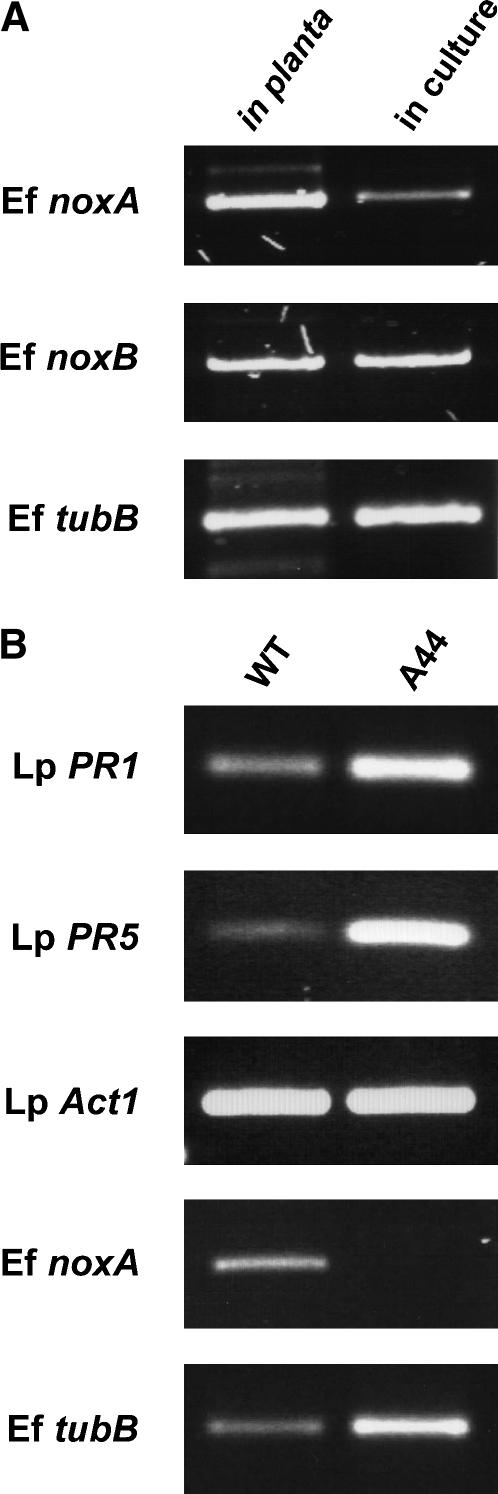
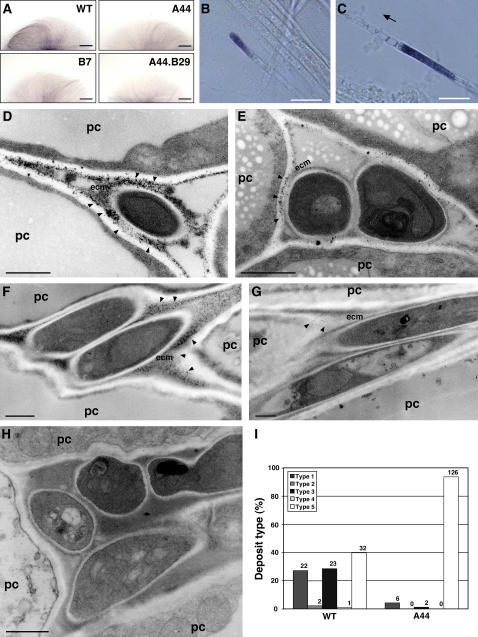
Although much is known about the signals and mechanisms that lead to pathogenic interactions between plants and fungi, comparatively little is known about fungus-plant mutualistic symbioses. We describe a novel role for reactive oxygen species (ROS) in regulating the mutualistic interaction between a clavicipitaceous fungal endophyte, Epichloë festucae, and its grass host, Lolium perenne. In wild-type associations, E. festucae grows systemically in intercellular spaces of leaves as infrequently branched hyphae parallel to the leaf axis. A screen to identify symbiotic genes isolated a fungal mutant that altered the interaction from mutualistic to antagonistic. This mutant has a single-copy plasmid insertion in the coding region of a NADPH oxidase gene, noxA. Plants infected with the noxA mutant lose apical dominance, become severely stunted, show precocious senescence, and eventually die. The fungal biomass in these associations is increased dramatically, with hyphae showing increased vacuolation. Deletion of a second NADPH oxidase gene, noxB, had no effect on the E. festucae-perennial ryegrass symbiosis. ROS accumulation was detected cytochemically in the endophyte extracellular matrix and at the interface between the extracellular matrix and host cell walls of meristematic tissue in wild-type but not in noxA mutant associations. These results demonstrate that fungal ROS production is critical in maintaining a mutualistic fungus-plant interaction.
Figures
References
-
- Aguirre, J., Ríos-Momberg, M., Hewitt, D., and Hansberg, W. (2005). Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 13 111–118. - PubMed
-
- Akiyama, K., Matsuzaki, K., and Hayashi, H. (2005). Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435 824–827. - PubMed
-
- Bennett, J.W., and Lasure, L.L. (1985). Conventions for gene symbols. In Gene Manipulation in Fungi, J.W. Bennett and L.L. Lasure, eds (London: Academic Press), pp. 537–544.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
