

Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell-cell fusion but does not affect virion entry
- PMID: 16519916
- PMCID: PMC7111780
- DOI: 10.1016/j.virol.2006.02.003
Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell-cell fusion but does not affect virion entry
Abstract
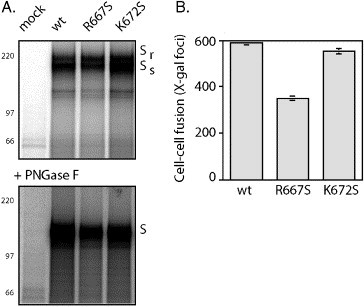
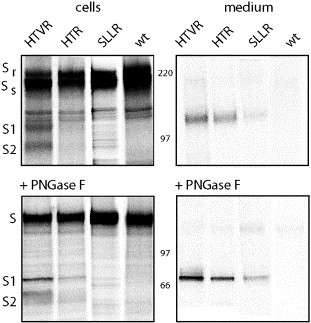
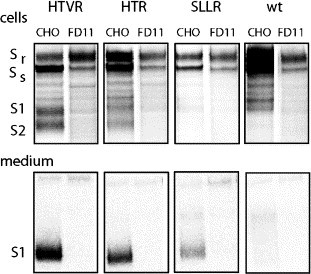
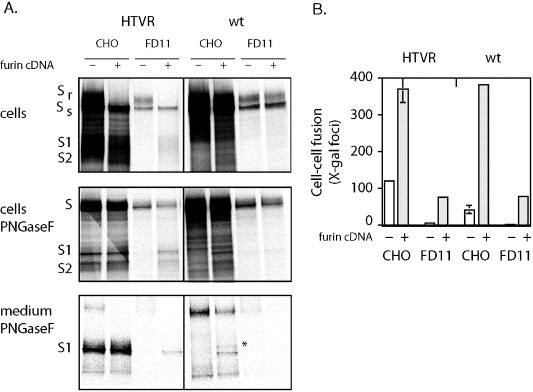
The fusogenic potential of Class I viral envelope glycoproteins is activated by proteloytic cleavage of the precursor glycoprotein to generate the mature receptor-binding and transmembrane fusion subunits. Although the coronavirus (CoV) S glycoproteins share membership in this class of envelope glycoproteins, cleavage to generate the respective S1 and S2 subunits appears absent in a subset of CoV species, including that responsible for the severe acute respiratory syndrome (SARS). To determine whether proteolytic cleavage of the S glycoprotein might be important for the newly emerged SARS-CoV, we introduced a furin recognition site at single basic residues within the putative S1-S2 junctional region. We show that furin cleavage at the modified R667 position generates discrete S1 and S2 subunits and potentiates membrane fusion activity. This effect on the cell-cell fusion activity by the S glycoprotein is not, however, reflected in the infectivity of pseudotyped lentiviruses bearing the cleaved glycoprotein. The lack of effect of furin cleavage on virion infectivity mirrors that observed in the normally cleaved S glycoprotein of the murine coronavirus and highlights an additional level of complexity in coronavirus entry.
Figures

References
-
- Berry J.D., Jones S., Drebot M.A., Andonov A., Sabara M., Yuan X.Y., Weingartl H., Fernando L., Marszal P., Gren J., Nicolas B., Andonova M., Ranada F., Gubbins M.J., Ball T.B., Kitching P., Li Y., Kabani A., Plummer F. Development and characterisation of neutralising monoclonal antibody to the SARS-coronavirus. J. Virol. Methods. 2004;120:87–96. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
