

Review
doi: 10.1021/cr050247k.
Hyaluronidases: their genomics, structures, and mechanisms of action
Affiliations
- PMID: 16522010
- PMCID: PMC2547145
- DOI: 10.1021/cr050247k
Item in Clipboard
Review
Hyaluronidases: their genomics, structures, and mechanisms of action
Chem Rev.
2006 Mar.
No abstract available
Figures
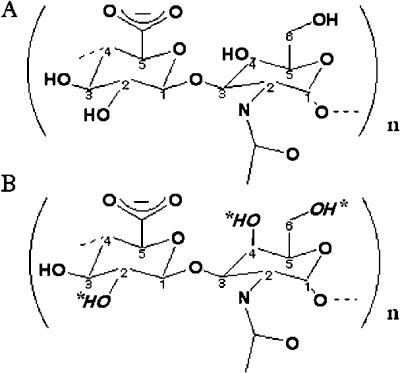
The substrate structures differ only in the anomericity at the C4 position of the N-acetyl-D-glycosamine, N-acetyl-D-glucosamine in hyaluronan and N-acetyl-D-galactosamine in chondroitin. Potential sulfation sites within the chondroitin molecule are indicated by italicized hydroxyl groups and asterisks. Both glycans are substrates for the human hyaluronidase enzymes. For Hyal-1 and -2, HA is the predominant substrate. However, binding and degradation of Ch/ChS also occurs, albeit at a slower rate, as observed experimentally in vitro. Hyal-4 appears to be a chondroitinase, with high specificity for Ch and ChS.
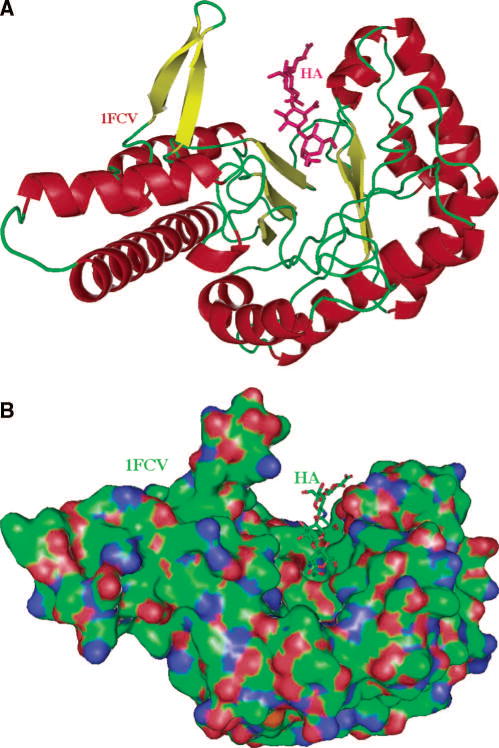
The only experimental structure of Hyal available is that of the bee venom enzyme. The historic secondary structure of Hyal is that of a homology model for BPH-20 and its complex with HA , which are similar to that of the native X-ray crystal structure of bee venom hyaluronidase and its complex with HA (pdb code: 1FCQ and 1FCV, respectively) . (A) The overall distorted (β/α)8 TIM barrel fold and the cleft are shown. The fold of this globular protein is characteristic of all glycoside hydrolases of carbohydrate active enzyme (CAZy) family 56. The molecule (based on coordinates of BVHyal having the pdb code: 1FCV) is color-coded by the secondary structure elements (helices in red, β-sheets in yellow, others in green). The HA molecule bound to the enzyme is located in the HA-binding cleft and is depicted in ball and stick fashion, and is colored in purple. This figure and other structural figures in this work were made with PyMol . (B) Surface of the molecule and a tetrasaccharide HA substrate bound within the cleft. The orientation of BVHyal molecule is similar to that in panel A. The protein surface is colored by the atomic element (C – green, N - blue, O - red, S - yellow). A large and positive charge and the hydrophobic character of the surface of the cleft allows for binding of negatively charged and also hydrophobic substrates. The cleft is where the catalytic function of hyaluronidases is performed. The HA tetrasaccharide is shown bound to the enzyme, also color coded by the atomic element.
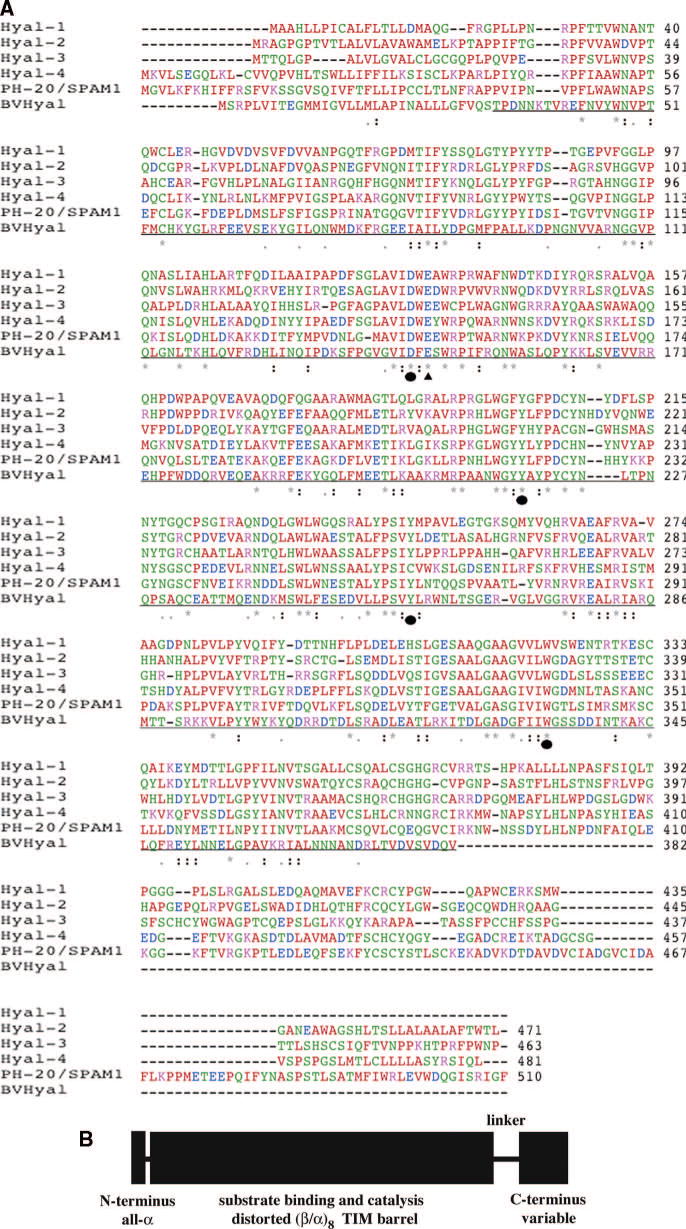
(A) Sequence alignment of the human hyaluronidases Hyal-1-Hyal-4, PH-20/SPAM1 based on the known X-ray structure of bee venom Hyal. The identity between all sequences varies from 33.1% between Hyal-3 and -4 to 41.2% between Hyal-4 and HPH-20 ,. The identity of the sequence of the BVHyal enzyme in the aligned region ranges from 22.9% for PH-20 to 25.2% for Hyal-1. The catalytic Glu H-donor residue and the residues positioning the nucleophile/base of the substrate are strictly conserved (Table 1), with the exception of Cys264 residue of Hyal-4, which may reflect specificity of this enzyme for Ch/ChS. The types of amino acid residues are color coded as follows: red - AVFPMILW small residue, green - STYHCNGQ hydroxyl, amine, or basic, blue - DE acidic, magenta - RK basic, and gray = others. The conserved residues are marked with ‘*’ – identical in entire column, ‘:’ – conserved according to color scheme above, and ‘.’ – semi-conserved substitutions are observed. The proposed catalytic Glu H-donating residue is in addition marked with ‘σ‘ whereas residues positioning the carbonyl nucleophile/base is marked with ‘λ‘. The sequence of the portion of the BVHyal enzyme that was crystallized is underlined. The sequences were aligned and the figure was made using Clastal W 1.82 . (B) Schematic diagram of domain composition of human Hyals. Human Hyals are composed of two domains, a major catalytic domain followed by a C-terminal one of unknown function. These domains are connected by a probably flexible peptide linker. The short segment at the extreme N-terminus is independent of the catalytic domain and assumes an α-helical conformation . The secondary structure elements for each domain are also indicated.
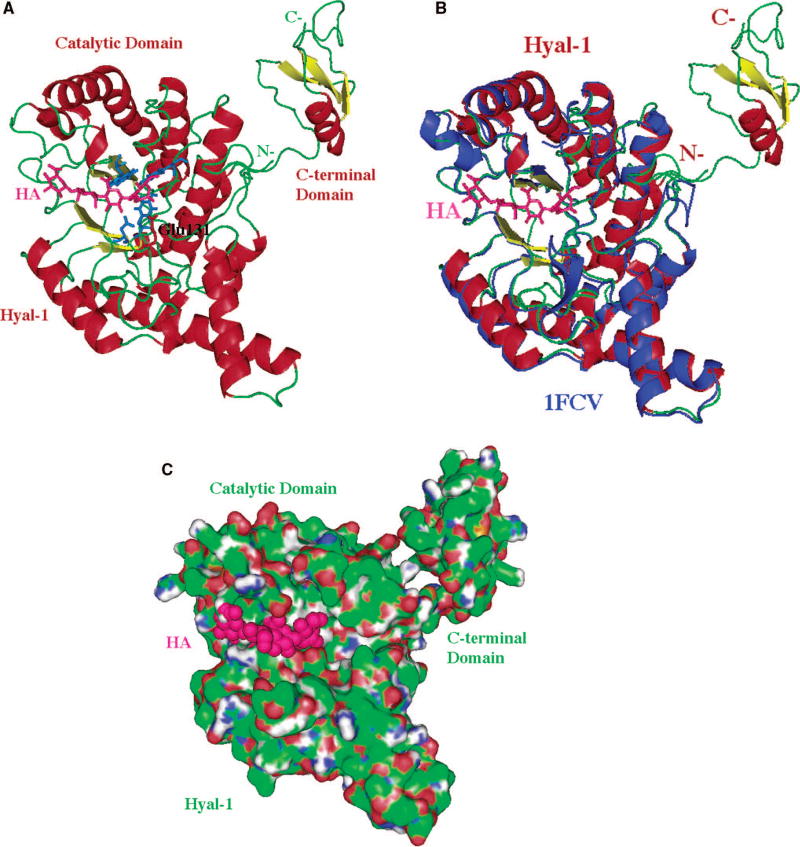
The structure of the model is based primarily on the comparative/homology modeling (for the main, catalytic domain) utilizing the structure of bee venom hyaluronidase (pdb code: 1FCQ) as described by Jedrzejas and Stern . The modeled C-terminal domain was obtained using the ab initio method and its primary, secondary, or tertiary structure is not similar to any other molecule’s sequence of 3D domain structure currently available, which implies a novel fold and possibly novel function. A characteristic (β/α)8 TIM barrel fold of the main domain, which is the catalytic domain that supports the binding to the substrate, and its hydrolytic degradation by means of double-displacement, retaining mechanism (described in the text). Reprinted with permission from Reference . Copyright 2005 John Wiley & Sons, Inc. (A) Three-dimensional model of Hyal-1 with bound hyaluronan. The molecule is color-coded by the secondary structure elements (same as in Fig. 2A). The bound HA molecule is located in the HA-binding cleft and is depicted in ball and stick fashion colored in purple. The catalytic residue, Glu131 is also shown in ball and stick fashion colored in red (labeled); other residues that position the carbonyl of the acetylamido group of HA are also shown and are colored in blue (not labeled). The catalytic and C-terminal domains, as well as the N- and C-termini are shown and labeled. (B) Comparison of 3D structure of Hyal-1 and BVHyl enzymes. The Hyal-1 molecule is shown in the same color and orientation as in panel A (including HA). The BVHyal enzyme structure is overlaid on the structure of the Hyal-1 model and colored in blue. The positions of the catalytic Glu and carbonyl positioning residues are essentially identical in the two structures (data not shown). The BVHyal does not have a C-terminal domain. (C) Surface of the Hyal-1 molecule and a tetrasaccharide HA substrate bound in the cleft. The orientation of the molecule is similar to that in panels A-B. The protein surface is colored as in Fig. 2B. The surface of the HA tetrasaccharide bound to the enzyme is shown and color in purple.
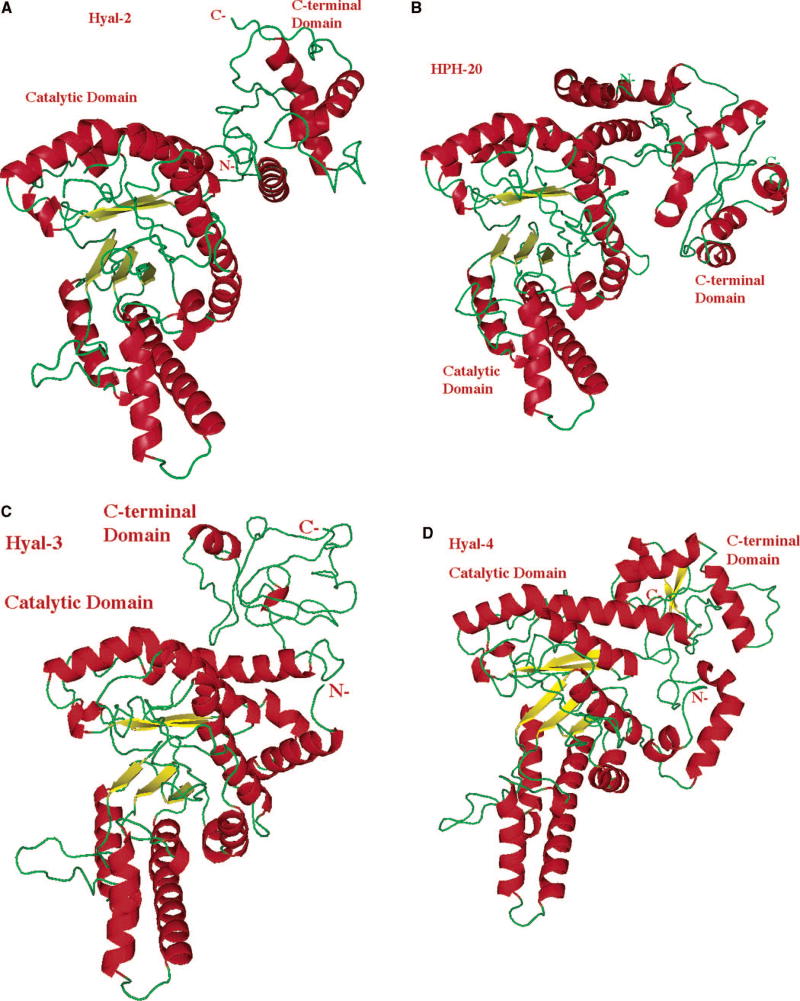
The molecule is sown in similar orientation and using the same color scheme and labeling as that of Hyal-1 in Fig. 4 A-B. The structures of the catalytic domains are similar to the same domain of Hyal-1 and to BVHyal enzyme. The structure and the size of the C-terminal domain are different between every Hyal and different from the same domain of Hyal-1. Reprinted with permission from Reference . Copyright 2005 John Wiley & Sons, Inc. (A) Model of Hyal-2, which degrades hyaluronan in human tissues to the approximate size of 50 disaccharides (~20 kDa). (B) Structural model of human PH-20/SPAM1 hyaluronidase found in sperm, involved in fertilization, as well as facilitating the penetration of sperm through the cumulus mass to reach the ovum. (C) Structural model of a relatively little investigated Hyal-3. (D) Model of Hyal-4 with the putative chondroitinase activity.
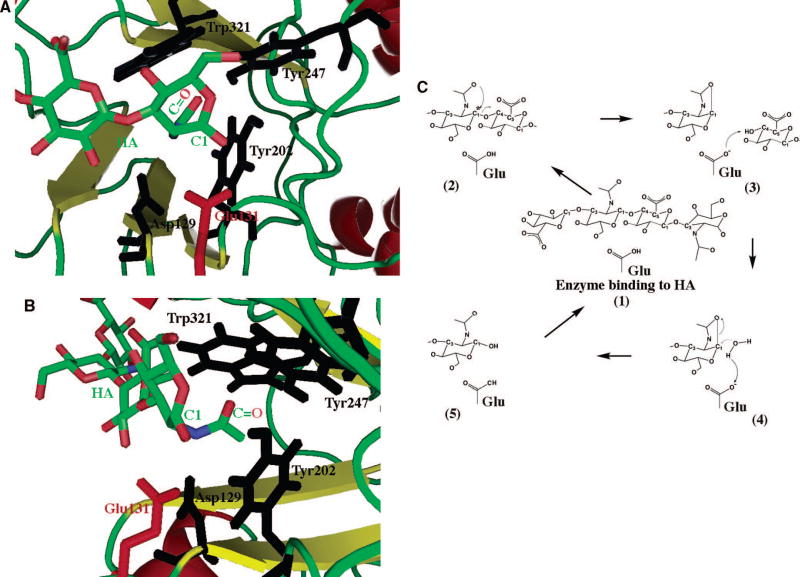
The scheme is based on the structure of Hyal-1 enzyme (based on structure of Hyal-1 as reported by Jedrzejas and Stern 12) with HA tetrasaccharide positions as in the structure of BVHyal homolog (pad coordinate code: 1FCV). The reducing end of bound HA molecule is located in the HA-binding cleft and is depicted in ball and stick fashion colored in by atom type as in Fig. 2B. The catalytic residue, Glu131 is also shown in ball and stick fashion colored in red (labeled); other residues that position the carbonyl of the acetylamido group (labeled) of HA are also shown and are colored in black (labeled). The C1 atom and carbonyl group (marked as C=O) of HA’s N-acetyl-D-glucosamine are labeled. (A) View from above HA and HA-binding cleft down towards the enzyme’s catalytic domain. The catalytic H-donor (acid) Glu131, in red, is shown in position to interact with the C1 carbon. The carbonyl positioning residues, in blue, are deeper in the enzyme’s cleft in which the acetamido group is located. (B) Alternate view from down the HA chain along the HA binding cleft. The carbonyl positioning residues are clearly in position to modify the position of the acetamido group to allow interaction of the carbonyl group with C1 carbon. (C) Schematic double displacement, retaining mechanism common for human hyaluronidases. This mechanism is characteristic of polysaccharide hydrolases and it involves the five depicted steps, 1–5, and one aa residue, Glu, of the enzyme (as described in the text). It involves a double displacement mechanism with the retention of C1 carbon (marked with an asterisks) configuration (retaining enzyme) . The catalytic process proceeds through an intermediate reaction step involving oxocarbonium ion at the C1 position of N-acetyl-D-glucosamine of the substrate. The catalytic acid function is performed by an absolutely conserved mechanism among Hyal enzymes Glu residue. The nucleophile/base function is attributes to the carbonyl oxygen of C-2 acetamido group of the substrate: HA, Ch, or ChS.
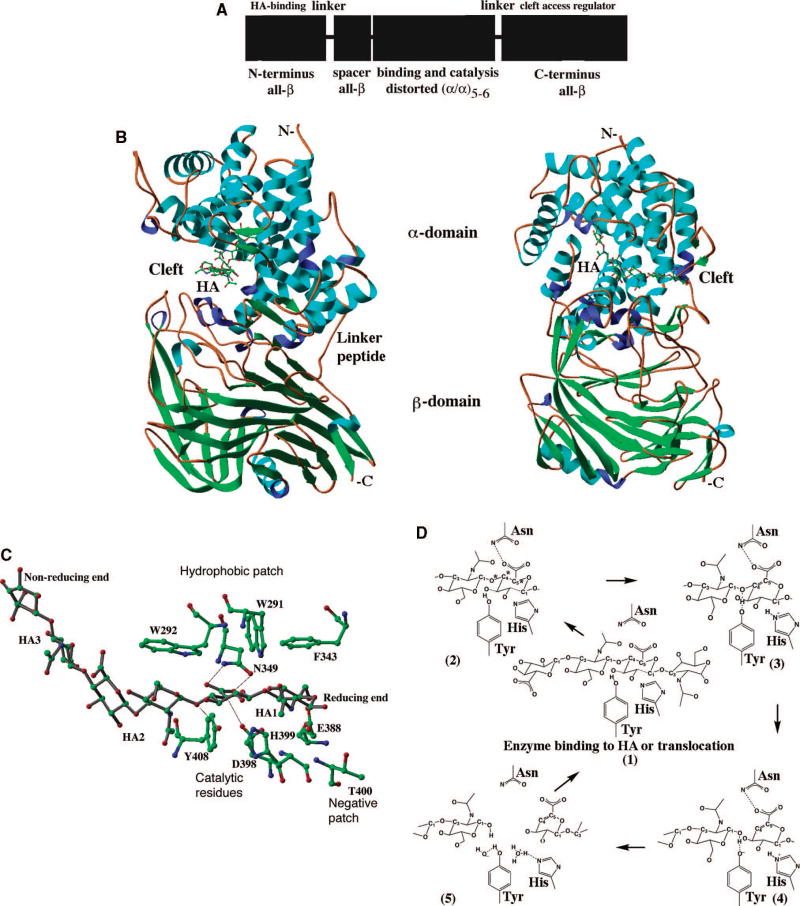
(A) Schematic diagram of domain composition of bacterial Hyals. Bacterial Hyals are composed of four domains, a HA-binding module, a spacer domain, a major catalytic domain (α-domain), followed by a C-terminal one (β-domain) that regulates HA access to the cleft . These N-terminal HA-binding module and the most C-terminal domains are connected to the rest of the protein by presumably flexible peptide linkers. The module at the extreme N-terminus is independent of the rest of the protein and assumes an all β-sheet structure. The secondary structure elements for each domain are indicated. (B) Overall structure. The crystallized enzyme is built from an α-helical catalytic domain and a supportive β-sheet domain (based on structure coordinates, pdb code: 1LOH) . Both domains are connected by one flexible peptide linker. The most N-terminal two domains, a spacer domain (composed of β-sheets) and the additional HA-binding one (α/β fold) are included in this structure . Two perpendicular views of the enzyme (rotated along the axis parallel to the labeled N- - C-termini) are shown. The hexasaccharide hyaluronan substrate is depicted in the enzyme’s cleft in a ball and stick fashion color coded by atomic element as in Fig. 2B. The molecule is colored by the secondary structure elements (α-helices- blue, 3/10-helices- purple, β-sheets- green, coil regions- brown). N-and C-termini are labeled. (C) Catalytic residues of the enzyme. The residues directly involved in catalysis are Asn349, His399, and Tyr408 (catalytic group); positioning of the substrate are Trp291, Trp292, and Phe343 (hydrophobic patch); release of the product are Glu388, Asp398, and Thr400 (negative patch) are shown together with the hyaluronan hexasaccharide substrate (based on structure coordinates, pdb code: 1LOH) . The most essential enzyme-substrate interactions for the catalytic process are shown as black lines. Consecutive HA disacchrides from the reducing to non-reducing end are labeled HA1-HA3. All residues and HA hexasaccharide are shown in ball and stick fashion and color coded by atomic element (as in Fig. 2B). (D) Proton acceptance and donation mechanism of bacterial hyaluronan lyases. This mechanism consists of a five steps process and involved three residues of the enzyme, Asn, His, and Tyr, as described in the text ,. The glycosidic oxygen as well as C4 and C5 carbon atoms directly involved in catalysis are also marked by asterisks. There is no direct water molecule involvement in this catalytic process except during H exchange to ready enzyme for the next round of catalysis. Smaller HA chains are primarily degraded in a processive manner, whereas large aggregated HA molecules, Ch and ChS with selected sulfation patterns (as described in the text) of any length are degraded non-processively .
References
-
- Rigden DJ, Jedrzejas MJ. J Biol Chem. 2003;278:50596–606. - PubMed
-
- Jedrzejas MJ. Crit Rev Biochem Mol Biol. 2000;35:221–51. - PubMed
-
- Jedrzejas MJ. Front Biosci. 2004;9:891–914. - PubMed
-
- Jedrzejas MJ. In: Hyaluronan 2003. Hascall VC, Balazs EA, editors. Matrix Bilogy Institute; Edgewater, NJ: 2004. www.matrixbiologyinstitute.org/ha03.
-
- Jedrzejas MJ. In: Recent Research Developments in Biophysics and Biochemistry. Pandali SG, editor. Vol. 2 Research Signpost; Trivandrum, India: 2002.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
