

Human replication protein A can suppress the intrinsic in vitro mutator phenotype of human DNA polymerase lambda
- PMID: 16522650
- PMCID: PMC1390690
- DOI: 10.1093/nar/gkl032
Human replication protein A can suppress the intrinsic in vitro mutator phenotype of human DNA polymerase lambda
Abstract
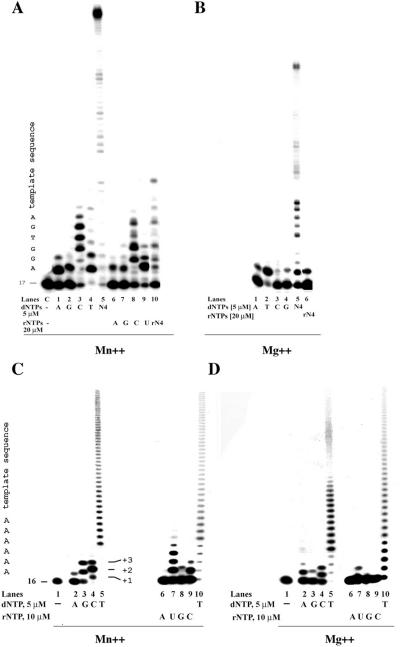
DNA polymerase lambda (pol lambda) is a member of the X family DNA polymerases and is endowed with multiple enzymatic activities. In this work we investigated the in vitro miscoding properties of full-length, human pol lambda either in the absence or in the presence of the human auxiliary proteins proliferating cell nuclear antigen (PCNA) and replication protein A (RP-A). Our data suggested that (i) pol lambda had an intrinsic ability to create mismatches and to incorporate ribonucleotides at nearly physiological Mn++ and Mg++ concentrations; (ii) the sequence of the template-primer could influence the misincorporation frequency of pol lambda; (iii) pol lambda preferentially generated G:T and G:G mismatches; (iv) RP-A, but not PCNA, selectively prevented misincorporation of an incorrect nucleotide by pol lambda, without affecting correct incorporation and (v) this inhibitory effect required a precise ratio between the concentrations of pol lambda and RP-A. Possible physiological implications of these findings for the in vivo fidelity of pol lambda are discussed.
Figures




References
-
- Hubscher U., Maga G., Spadari S. Eukaryotic DNA polymerases. Annu Rev. Biochem. 2002;71:133–163. - PubMed
-
- Garcia-Diaz M., Dominguez O., Lopez-Fernandez L.A., de Lera L.T., Saniger M.L., Ruiz J.F., Parraga M., Garcia-Ortiz M.J., Kirchhoff T., del Mazo J., et al. DNA polymerase lambda (Pol lambda), a novel eukaryotic DNA polymerase with a potential role in meiosis. J. Mol. Biol. 2000;301:851–867. - PubMed
-
- Ramadan K., Shevelev I., Hubscher U. The DNA-polymerase-X family: controllers of DNA quality? Nat. Rev. Mol. Cell Biol. 2004;5:1038–1043. - PubMed
-
- Maga G., Villani G., Ramadan K., Shevelev I., Le Gac N.T., Blanco L., Blanca G., Spadari S., Hubscher U. Human DNA polymerase lambda functionally and physically interacts with proliferating cell nuclear antigen in normal and translesion DNA synthesis. J. Biol. Chem. 2002;277:48434–48440. - PubMed
-
- Maga G., Blanca G., Shevelev I., Frouin I., Ramadan K., Spadari S., Villani G., Hubscher U. The human DNA polymerase lambda interacts with PCNA through a domain important for DNA primer binding and the interaction is inhibited by p21/WAF1/CIP1. Faseb J. 2004;18:1743–1745. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
