

Novel subcellular distribution pattern of A-type K+ channels on neuronal surface
- PMID: 16525047
- PMCID: PMC1558001
- DOI: 10.1523/JNEUROSCI.5257-05.2006
Novel subcellular distribution pattern of A-type K+ channels on neuronal surface
Abstract
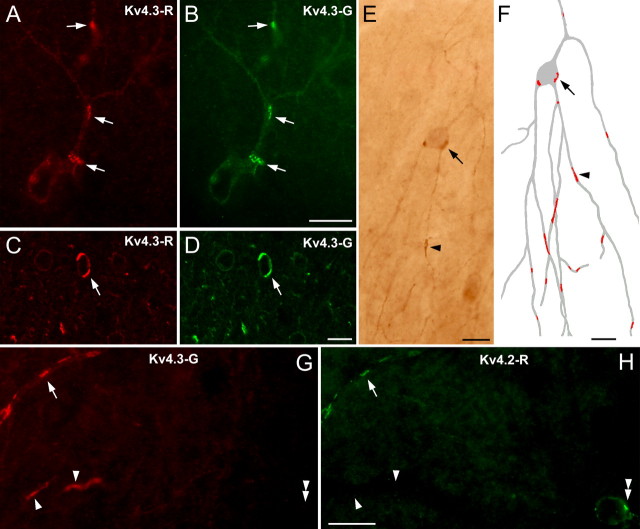
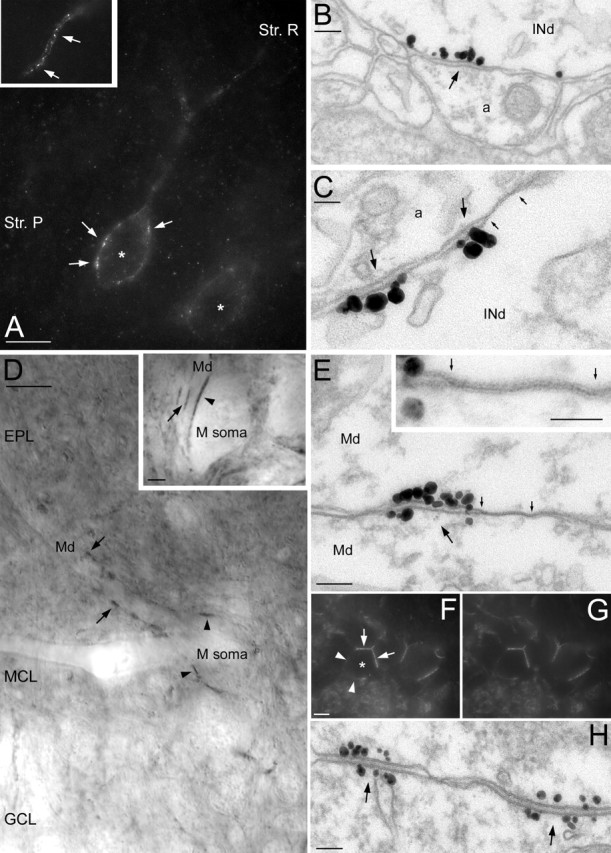
Potassium channels comprise the most diverse family of ion channels. In nerve cells, their critical roles in synaptic integration and output generation have been demonstrated. Here, we provide evidence for a distribution that predicts a novel role of K+ channels in the CNS. Our experiments revealed a highly selective clustering of the Kv4.3 A-type K+ channel subunits at specialized junctions between climbing fibers and cerebellar GABAergic interneurons. High-resolution ultrastructural and immunohistochemical experiments demonstrated that these junctions are distinct from known chemical and electrical (gap junctions) synapses and also from puncta adherentia. Each cerebellar interneuron contains many such K+ channel-rich specializations, which seem to be distributed throughout the somatodendritic surface. We also show that such K+ channel-rich specializations are not only present in the cerebellum but are widespread in the rat CNS. For example, mitral cells of the main olfactory bulb establish Kv4.2 subunit-positive specializations with each other. At these specializations, both apposing membranes have a high density of K+ channels, indicating bidirectional signaling. Similar specializations with pronounced coclustering of the Kv4.2 and 4.3 subunits were observed between nerve cells in the medial nucleus of the habenula. Based on our results and on the known properties of A-type K+ channels, we propose that strategically clustered K+ channels at unique membrane specializations could mediate a novel type of communication between nerve cells.
Figures




References
-
- Alonso G, Widmer H (1997). Clustering of Kv4.2 potassium channels in postsynaptic membrane of rat supraoptic neurons: an ultrastructural study. Neuroscience 77:617–621. - PubMed
-
- An WF, Bowlby MR, Betty M, Cao J, Ling HP, Mendoza G, Hinson JW, Mattsson KI, Strassle BW, Trimmer JS, Rhodes KJ (2000). Modulation of A-type potassium channels by a family of calcium sensors. Nature 403:553–556. - PubMed
-
- Cai X, Liang CW, Muralidharan S, Kao JP, Tang CM, Thompson SM (2004). Unique roles of SK and Kv4.2 potassium channels in dendritic integration. Neuron 44:351–364. - PubMed
-
- Chandy KG (1991). Simplified gene nomenclature. Nature 352:26. - PubMed
-
- Csiffary A, Gorcs TJ, Palkovits M (1990). Neuropeptide Y innervation of ACTH-immunoreactive neurons in the arcuate nucleus of rats: a correlated light and electron microscopic double immunolabeling study. Brain Res 506:215–222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous