

Depolarization of cochlear outer hair cells evokes active hair bundle motion by two mechanisms
- PMID: 16525055
- PMCID: PMC6675159
- DOI: 10.1523/JNEUROSCI.3808-05.2006
Depolarization of cochlear outer hair cells evokes active hair bundle motion by two mechanisms
Abstract
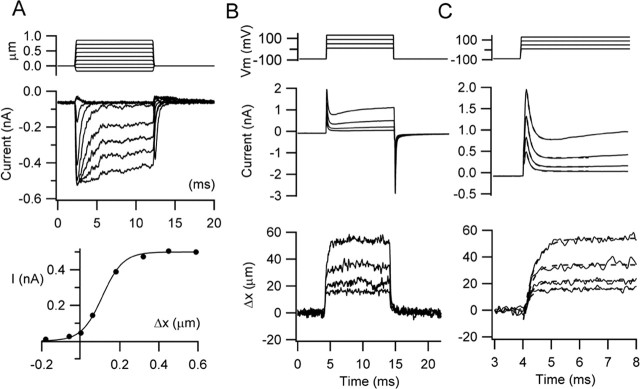
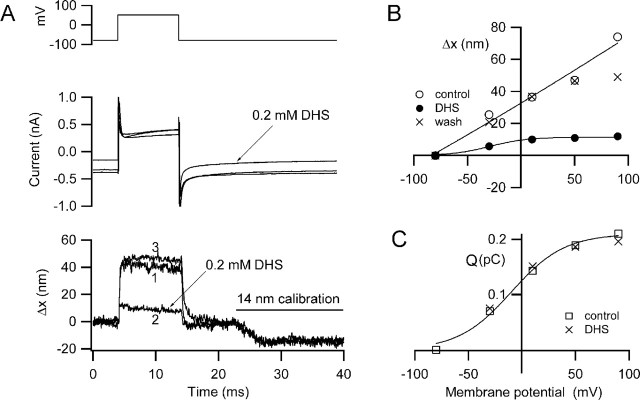
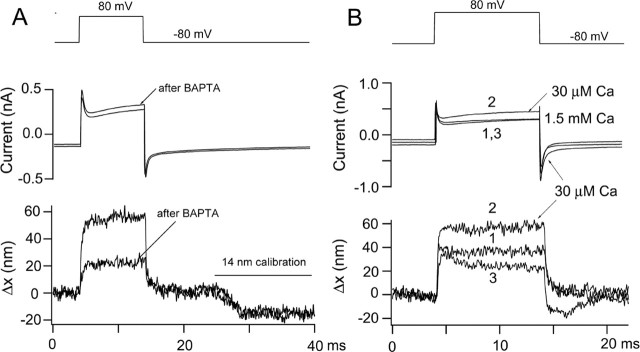
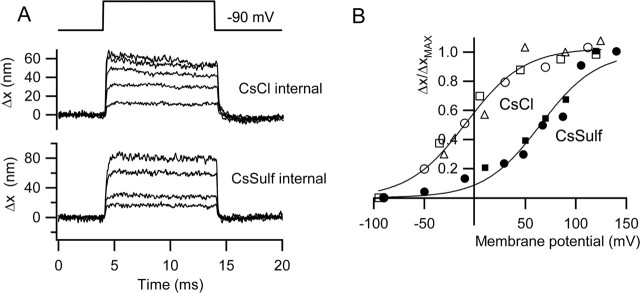
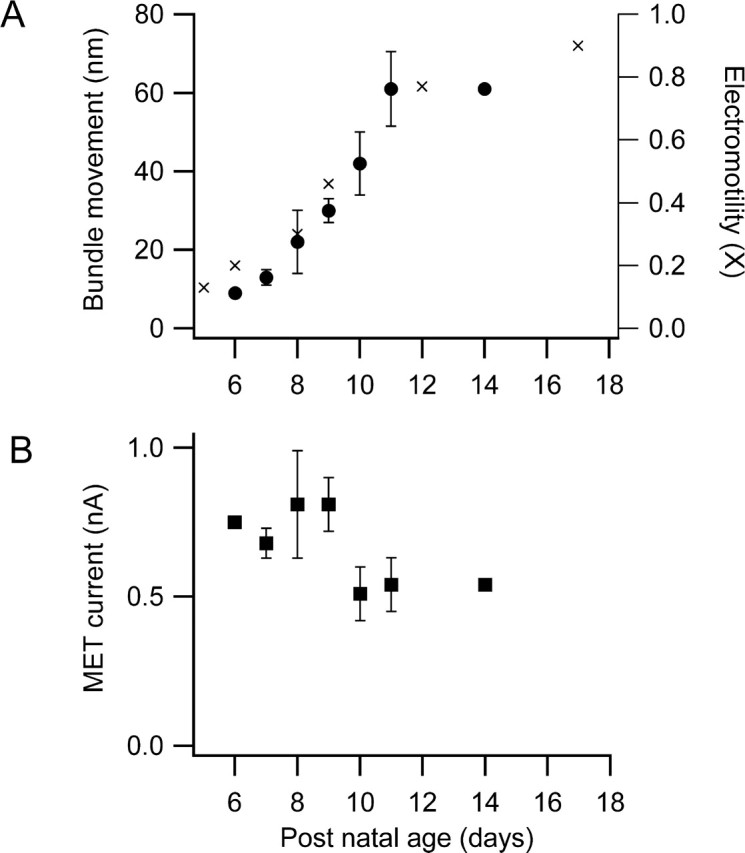
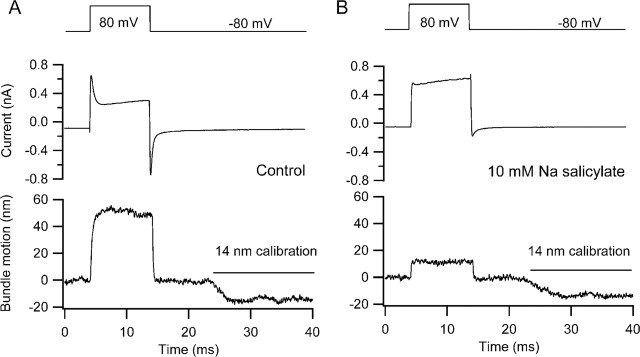
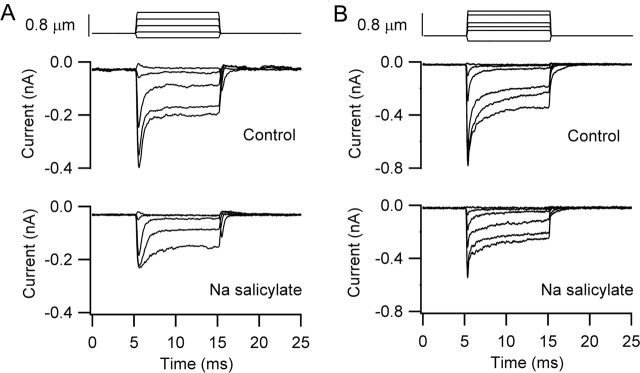
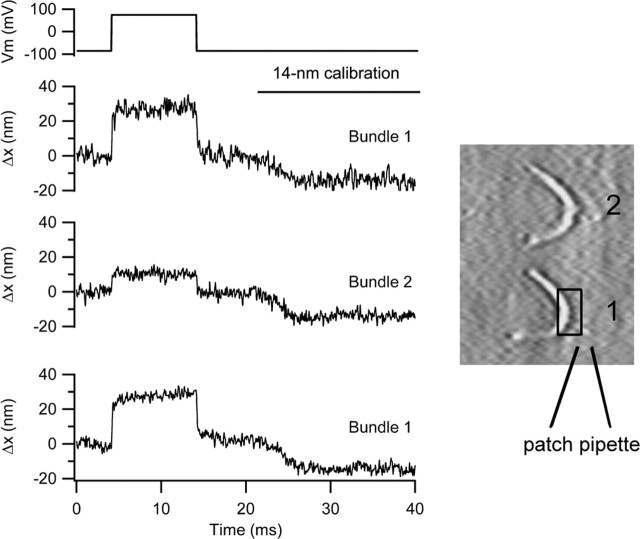
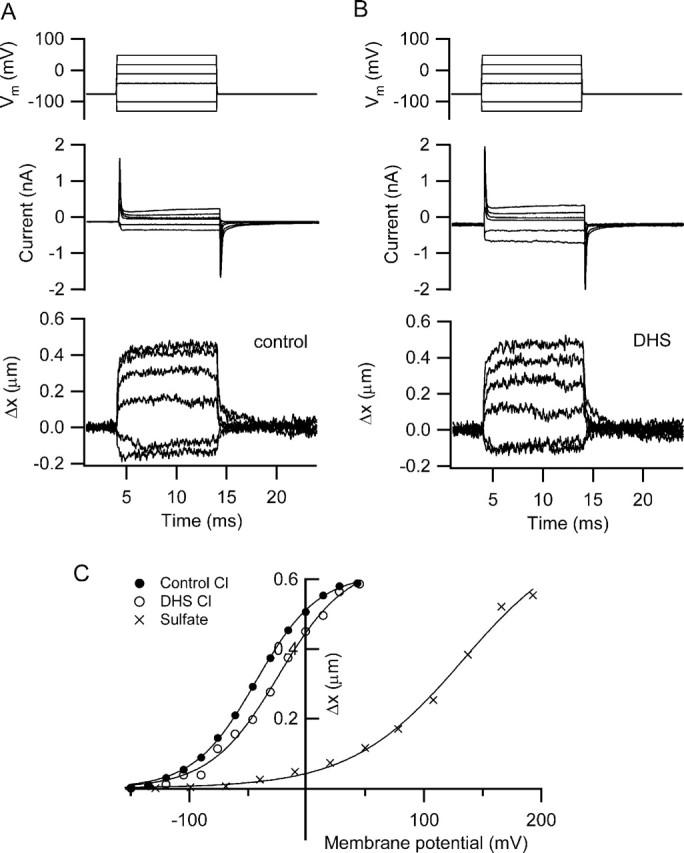
There is current debate about the origin of mechanical amplification whereby outer hair cells generate force to augment the sensitivity and frequency selectivity of the mammalian cochlea. To distinguish contributions to force production from the mechanotransducer (MET) channels and somatic motility, we have measured hair bundle motion during depolarization of individual outer hair cells in isolated rat cochleas. Depolarization evoked rapid positive bundle deflections that were reduced by perfusion with the MET channel blocker dihydrostreptomycin, with no effect on the nonlinear capacitance that is a manifestation of prestin-driven somatic motility. However, the movements were also diminished by Na salicylate and depended on the intracellular anion, properties implying involvement of the prestin motor. Furthermore, depolarization of one outer hair cell caused motion of neighboring hair bundles, indicating overall motion of the reticular lamina. Depolarization of solitary outer hair cells caused cell-length changes whose voltage-activation range depended on the intracellular anion but were insensitive to dihydrostreptomycin. These results imply that both the MET channels and the somatic motor participate in hair bundle motion evoked by depolarization. It is conceivable that the two processes can interact, a signal from the MET channels being capable of modulating the activity of the prestin motor.
Figures
Similar articles
-
Tmc1 Point Mutation Affects Ca2+ Sensitivity and Block by Dihydrostreptomycin of the Mechanoelectrical Transducer Current of Mouse Outer Hair Cells.J Neurosci. 2016 Jan 13;36(2):336-49. doi: 10.1523/JNEUROSCI.2439-15.2016. J Neurosci. 2016. PMID: 26758827 Free PMC article.
-
A prestin motor in chicken auditory hair cells: active force generation in a nonmammalian species.Neuron. 2013 Jul 10;79(1):69-81. doi: 10.1016/j.neuron.2013.05.018. Epub 2013 Jun 6. Neuron. 2013. PMID: 23746629 Free PMC article.
-
Cochlear amplification, outer hair cells and prestin.Curr Opin Neurobiol. 2008 Aug;18(4):370-6. doi: 10.1016/j.conb.2008.08.016. Epub 2008 Oct 4. Curr Opin Neurobiol. 2008. PMID: 18809494 Free PMC article. Review.
-
Active hair bundle movements in auditory hair cells.J Physiol. 2006 Oct 1;576(Pt 1):29-36. doi: 10.1113/jphysiol.2006.115949. Epub 2006 Aug 3. J Physiol. 2006. PMID: 16887874 Free PMC article. Review.
-
Motility-associated hair-bundle motion in mammalian outer hair cells.Nat Neurosci. 2005 Aug;8(8):1028-34. doi: 10.1038/nn1509. Epub 2005 Jul 24. Nat Neurosci. 2005. PMID: 16041370
Cited by
-
A clinically oriented introduction and review on finite element models of the human cochlea.Biomed Res Int. 2014;2014:975070. doi: 10.1155/2014/975070. Epub 2014 Nov 4. Biomed Res Int. 2014. PMID: 25530973 Free PMC article.
-
Coupling and elastic loading affect the active response by the inner ear hair cell bundles.PLoS One. 2012;7(3):e33862. doi: 10.1371/journal.pone.0033862. Epub 2012 Mar 27. PLoS One. 2012. PMID: 22479461 Free PMC article.
-
Energy Flux in the Cochlea: Evidence Against Power Amplification of the Traveling Wave.J Assoc Res Otolaryngol. 2015 Oct;16(5):581-97. doi: 10.1007/s10162-015-0529-5. Epub 2015 Jul 7. J Assoc Res Otolaryngol. 2015. PMID: 26148491 Free PMC article.
-
Prestin's anion transport and voltage-sensing capabilities are independent.Biophys J. 2009 Apr 22;96(8):3179-86. doi: 10.1016/j.bpj.2008.12.3948. Biophys J. 2009. PMID: 19383462 Free PMC article.
-
Effects of cochlear loading on the motility of active outer hair cells.Proc Natl Acad Sci U S A. 2013 Apr 2;110(14):5474-9. doi: 10.1073/pnas.1302911110. Epub 2013 Mar 18. Proc Natl Acad Sci U S A. 2013. PMID: 23509256 Free PMC article.
References
-
- Ashmore JF (1989). Transducer motor coupling in cochlea outer hair cells. In: Cochlear mechanisms: structure, function and models (Wilson JP, Kemp DT, eds) , pp. 107–114. New York: Plenum.
-
- Assad JA, Shepherd GM, Corey DP (1991). Tip-link integrity and mechanical transduction in vertebrate hair cells. Neuron 7:985–994. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous