

Ligands for clathrin-mediated endocytosis are differentially sorted into distinct populations of early endosomes
- PMID: 16530046
- PMCID: PMC2660893
- DOI: 10.1016/j.cell.2005.12.038
Ligands for clathrin-mediated endocytosis are differentially sorted into distinct populations of early endosomes
Abstract
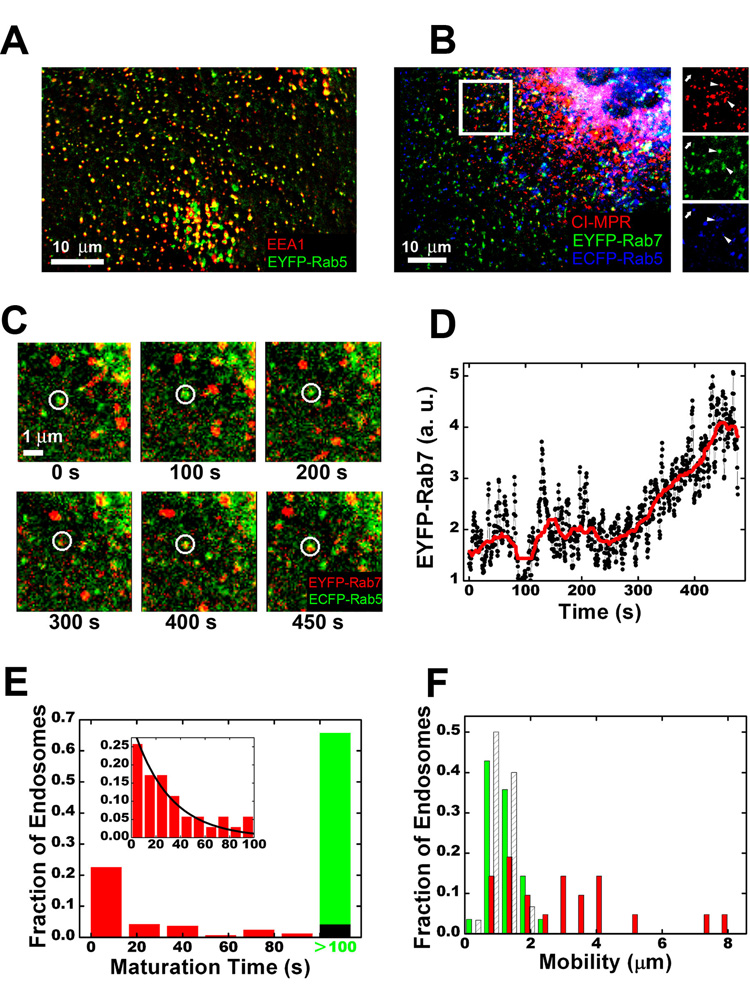
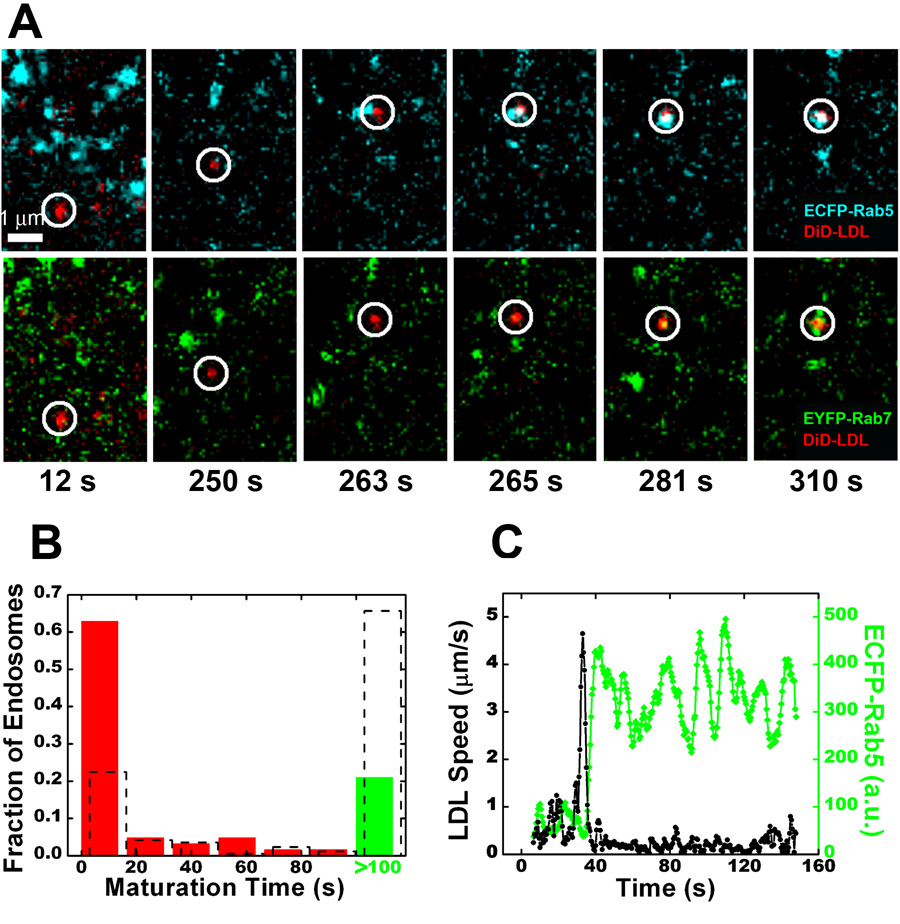
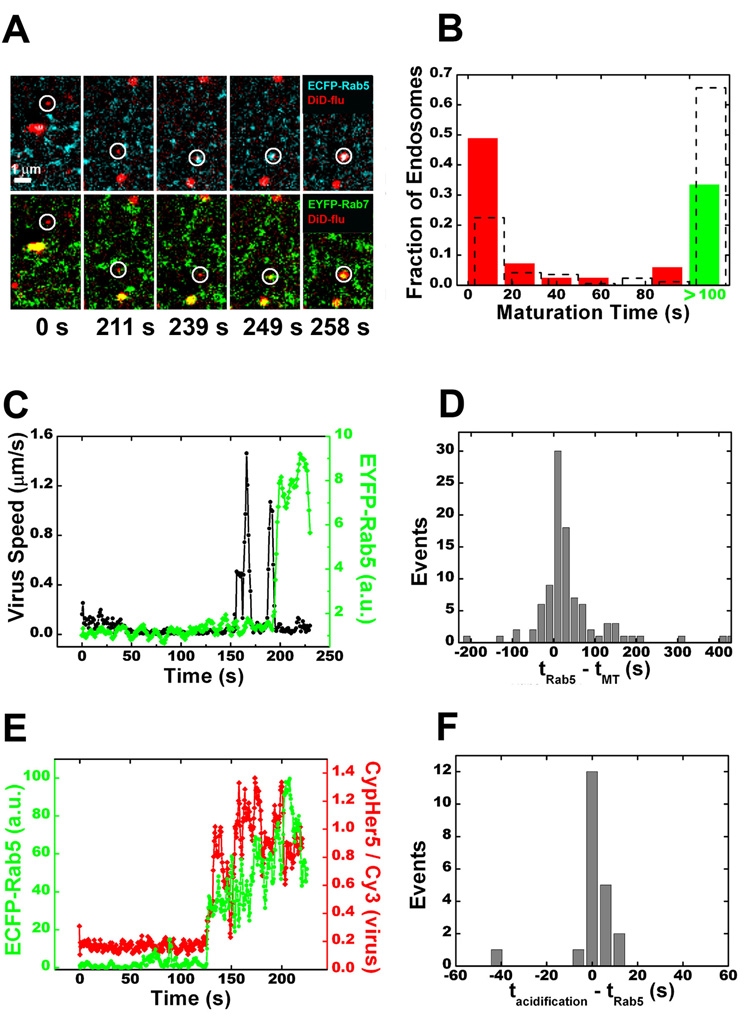
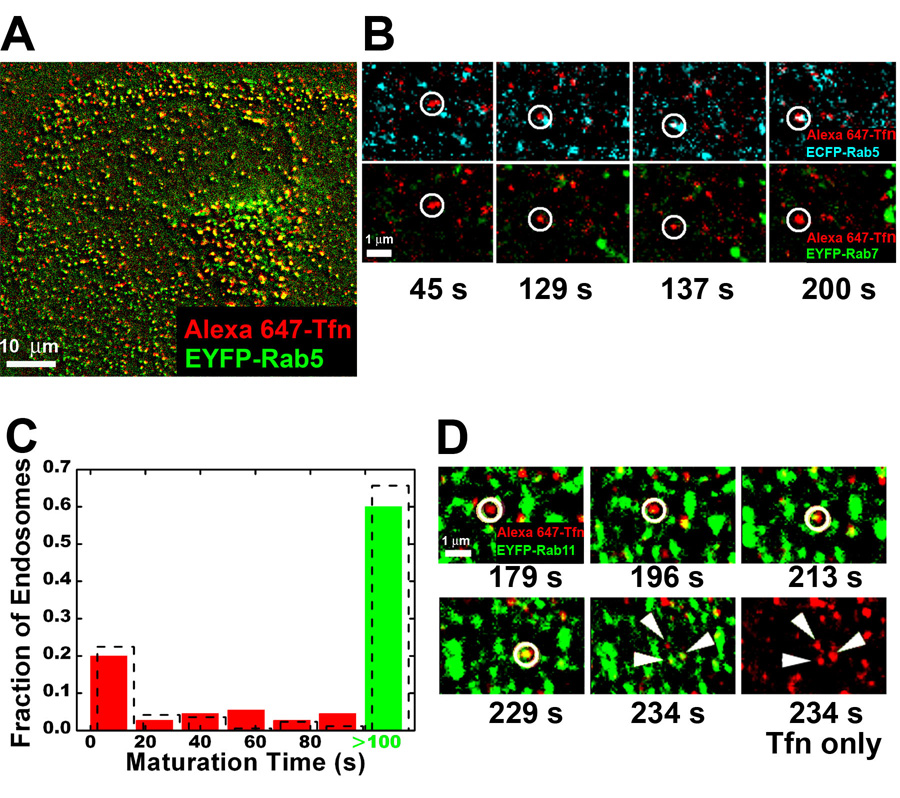
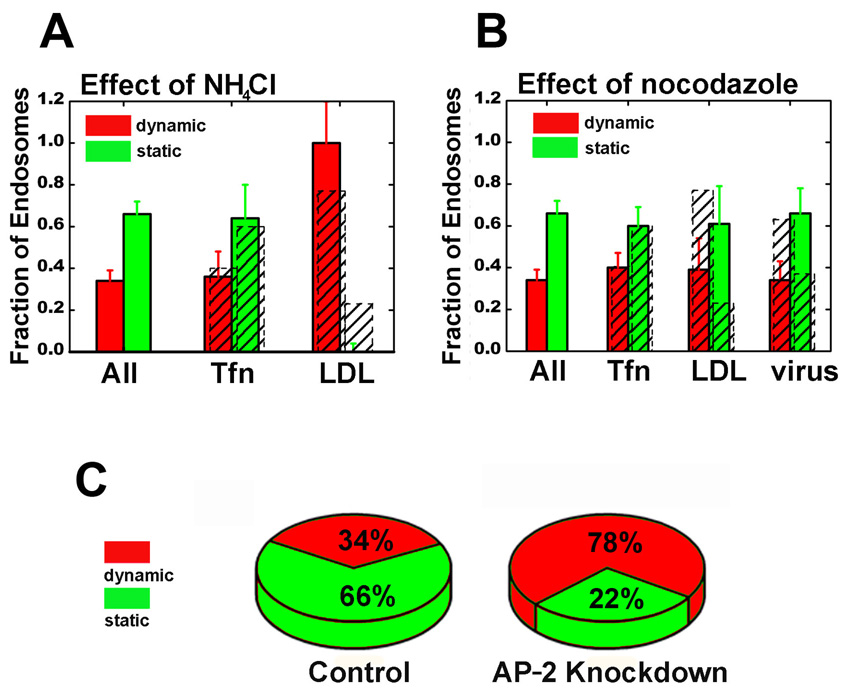
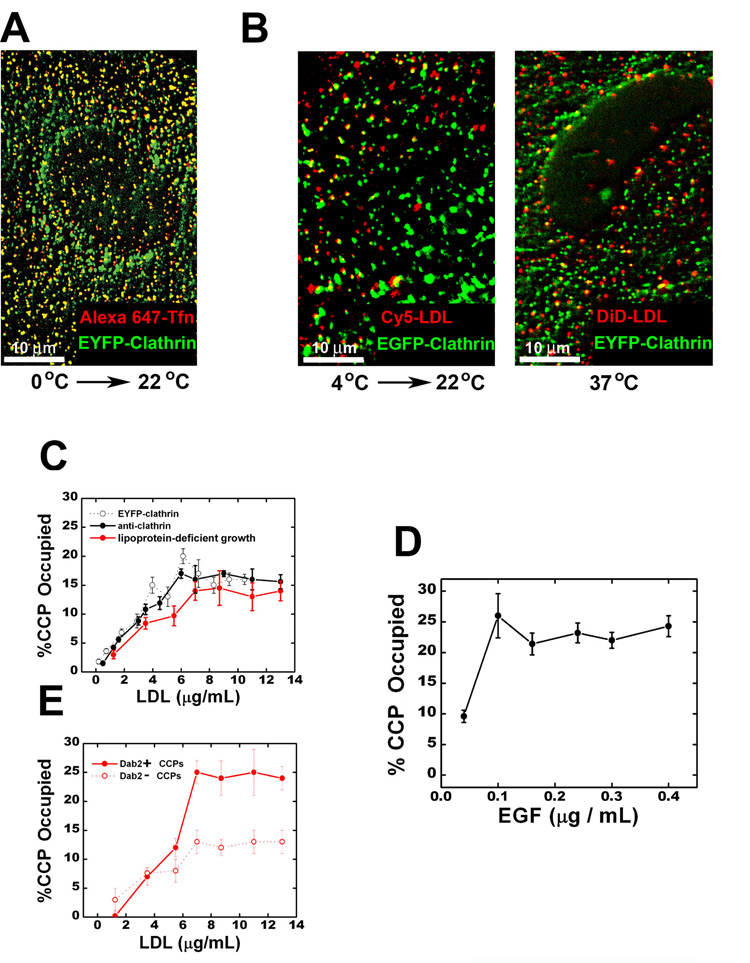
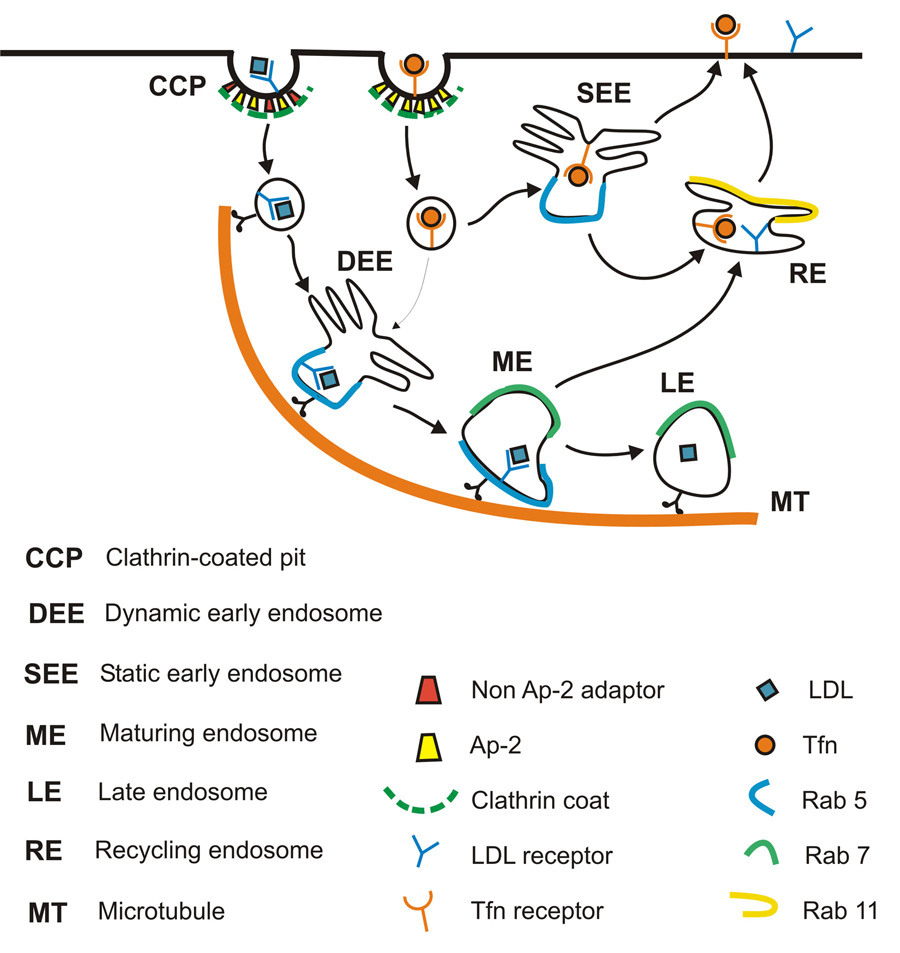
Cells rely on the correct sorting of endocytic ligands and receptors for proper function. Early endosomes have been considered as the initial sorting station where cargos for degradation separate from those for recycling. Using live-cell imaging to monitor individual endosomes and ligand particles in real time, we have discovered a sorting mechanism that takes place prior to early endosome entry. We show that early endosomes are in fact comprised of two distinct populations: a dynamic population that is highly mobile on microtubules and matures rapidly toward late endosomes and a static population that matures much more slowly. Several cargos destined for degradation are preferentially targeted to the dynamic endosomes, whereas the recycling ligand transferrin is nonselectively delivered to all early endosomes and effectively enriched in the larger, static population. This pre-early endosome sorting process begins at clathrin-coated vesicles, depends on microtubule-dependent motility, and appears to involve endocytic adaptors.
Figures
References
-
- Anderson RGW, Brown MS, Goldstein JL. Role of the coated endocytic vesicle in the uptake of receptor-bound low density lipoprotein in human fibroblasts. Cell. 1977;10:351–364. - PubMed
-
- Bonifacino JS, Lippincott-Schwartz J. Coated Proteins: shaping membrane transport. Nat Rev Mol Cell Biol. 2003;4:409–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
