

Conformational flexibility in the multidrug efflux system protein AcrA
- PMID: 16531241
- PMCID: PMC1997295
- DOI: 10.1016/j.str.2005.11.015
Conformational flexibility in the multidrug efflux system protein AcrA
Abstract
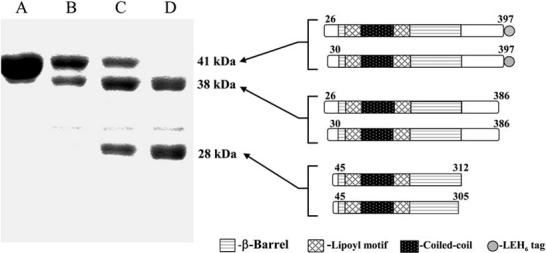
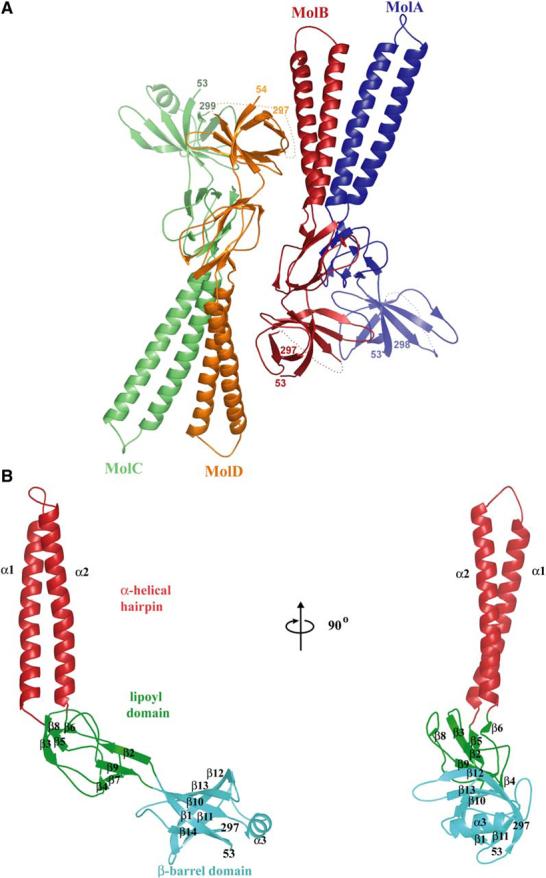
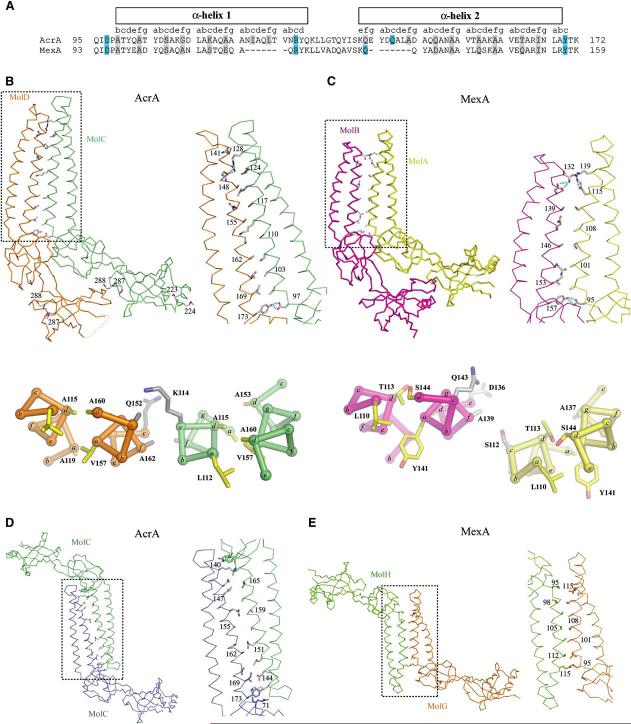
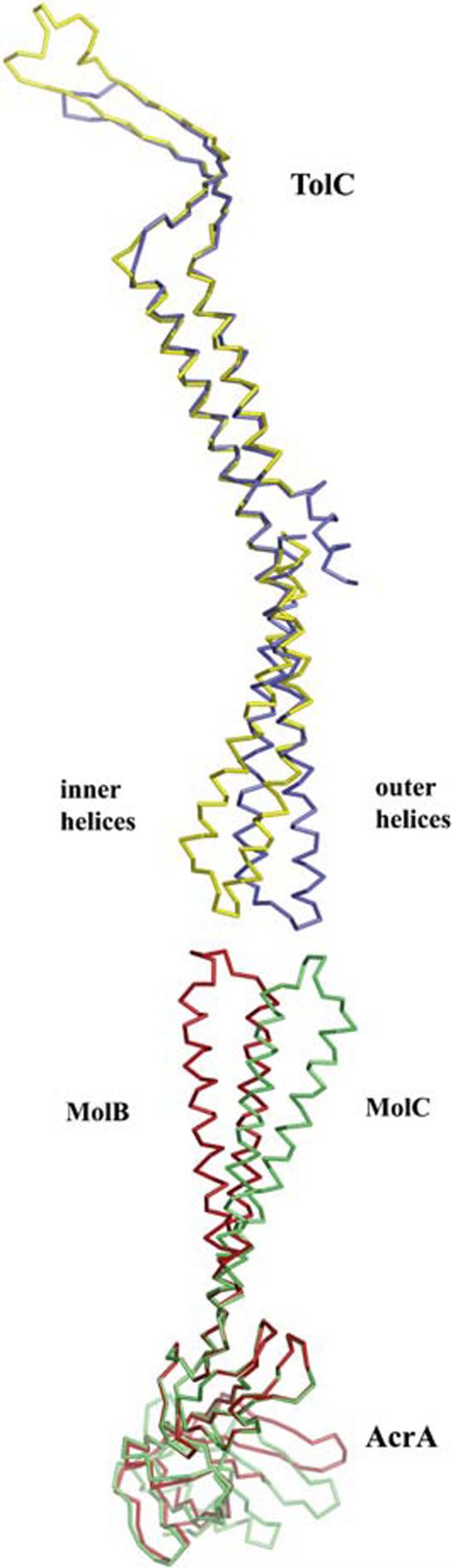
Intrinsic resistance to multiple drugs in many gram-negative bacterial pathogens is conferred by resistance nodulation cell division efflux pumps, which are composed of three essential components as typified by the extensively characterized Escherichia coli AcrA-AcrB-TolC system. The inner membrane drug:proton antiporter AcrB and the outer membrane channel TolC export chemically diverse compounds out of the bacterial cell, and require the activity of the third component, the periplasmic protein AcrA. The crystal structures of AcrB and TolC have previously been determined, and we complete the molecular picture of the efflux system by presenting the structure of a stable fragment of AcrA. The AcrA fragment resembles the elongated sickle shape of its homolog Pseudomonas aeruginosa MexA, being composed of three domains: beta-barrel, lipoyl, and alpha-helical hairpin. Notably, unsuspected conformational flexibility in the alpha-helical hairpin domain of AcrA is observed, which has potential mechanistic significance in coupling between AcrA conformations and TolC channel opening.
Figures


References
-
- Akama H, Kanemaki M, Yoshimura M, Tsukihara T, Kashiwagi T, Yoneyama H, Narita S, Nakagawa A, Nakae T. Crystal structure of the drug discharge outer membrane protein, OprM, of Pseudomonas aeruginosa: dual modes of membrane anchoring and occluded cavity end. J. Biol. Chem. 2004;279(a):52816–52819. - PubMed
-
- Akama H, Matsuura T, Kashiwagi S, Yoneyama H, Narita S, Tsukihara T, Nakagawa A, Nakae T. Crystal structure of the membrane fusion protein, MexA, of the multidrug transporter in Pseudomonas aeruginosa. J. Biol. Chem. 2004;279(b):25939–25942. - PubMed
-
- Budisa N, Steipe B, Demange P, Eckerskorn C, Kellermann J, Huber R. High-level biosynthetic substitution of methionine in proteins by its analogs 2-aminohexanoic acid, selenomethionine, telluromethionine and ethionine in Escherichia coli. Eur. J. Biochem. 1995;230:788–796. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
