

Xanthan induces plant susceptibility by suppressing callose deposition
- PMID: 16531487
- PMCID: PMC1459321
- DOI: 10.1104/pp.105.074542
Xanthan induces plant susceptibility by suppressing callose deposition
Abstract
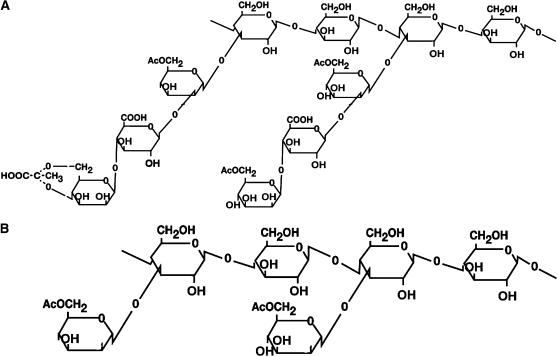
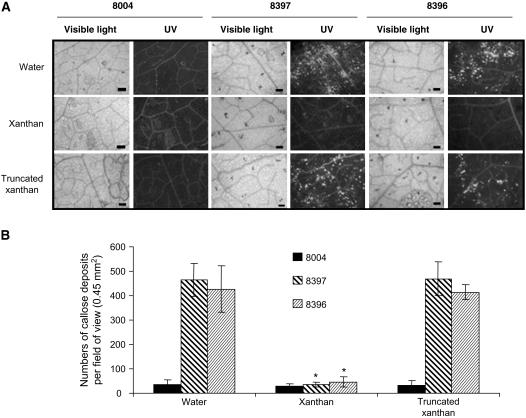
Xanthan is the major exopolysaccharide secreted by Xanthomonas spp. Despite its diverse roles in bacterial pathogenesis of plants, little is known about the real implication of this molecule in Xanthomonas pathogenesis. In this study we show that in contrast to Xanthomonas campestris pv campestris strain 8004 (wild type), the xanthan minus mutant (strain 8397) and the mutant strain 8396, which is producing truncated xanthan, fail to cause disease in both Nicotiana benthamiana and Arabidopsis (Arabidopsis thaliana) plants. In contrast to wild type, 8397 and 8396 strains induce callose deposition in N. benthamiana and Arabidopsis plants. Interestingly, treatment with xanthan but not truncated xanthan, suppresses the accumulation of callose and enhances the susceptibility of both N. benthamiana and Arabidopsis plants to 8397 and 8396 mutant strains. Finally, in concordance, we also show that treatment with an inhibitor of callose deposition previous to infection induces susceptibility to 8397 and 8396 strains. Thus, xanthan suppression effect on callose deposition seems to be important for Xanthomonas infectivity.
Figures





References
-
- Abramovitch RB, Martin GB (2004) Strategies used by bacterial pathogens to suppress plant defenses. Curr Opin Plant Biol 7: 356–364 - PubMed
-
- Abramovitch RB, Martin GB (2005) AvrPtoB: a bacterial type III effector that both elicits and suppresses programmed cell death associated with plant immunity. FEMS Microbiol Lett 245: 1–8 - PubMed
-
- Alfano JR, Collmer A (2004) Type III secretion system effector proteins: double agents in bacterial disease and plant defense. Annu Rev Phytopathol 42: 385–414 - PubMed
-
- Barrère GC, Barber CE, Daniels MJ (1986) Molecular cloning of genes involved in the production of the extracellular polysaccharide xanthan by Xanthomonas campestris pv campestris. Int J Biol Macromol 8: 372–374
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
