

Social isolation delays the positive effects of running on adult neurogenesis
- PMID: 16531997
- PMCID: PMC3029943
- DOI: 10.1038/nn1668
Social isolation delays the positive effects of running on adult neurogenesis
Abstract
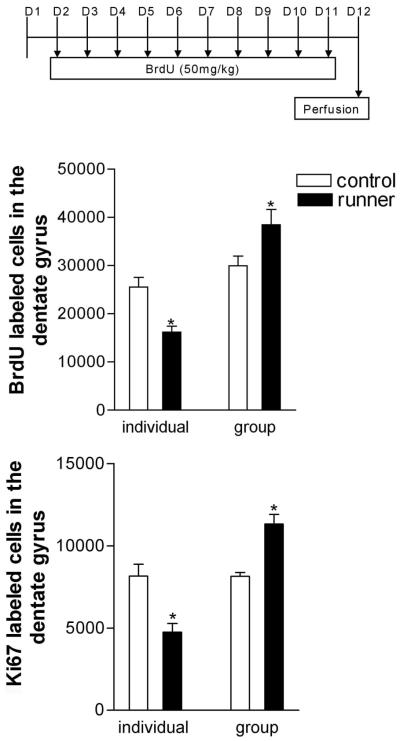
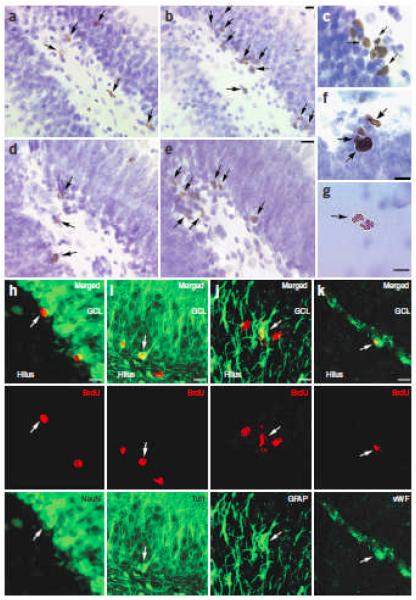
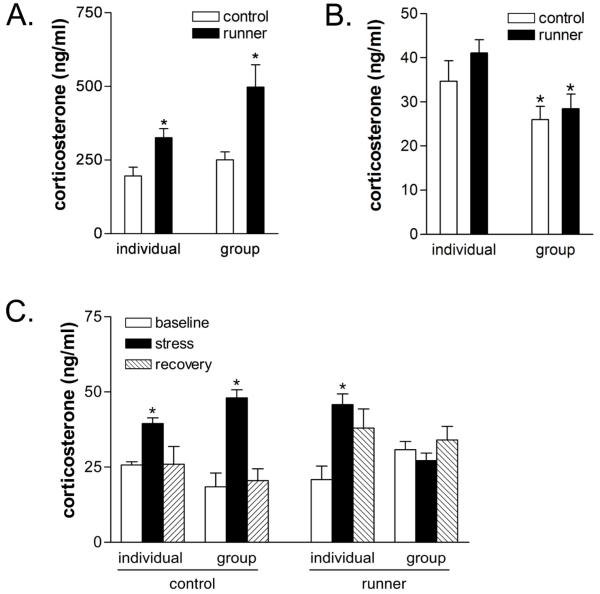
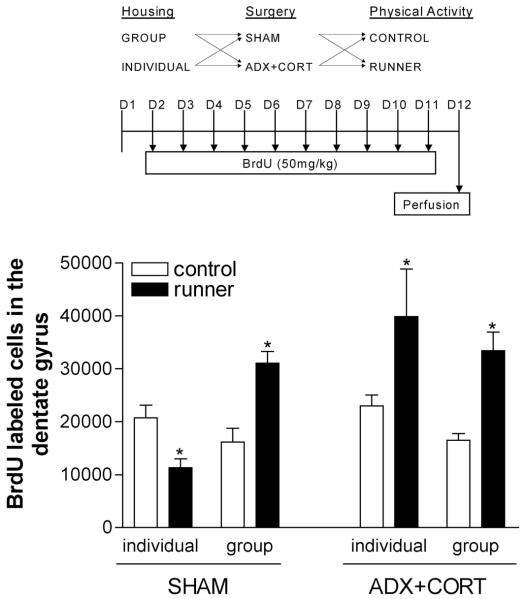
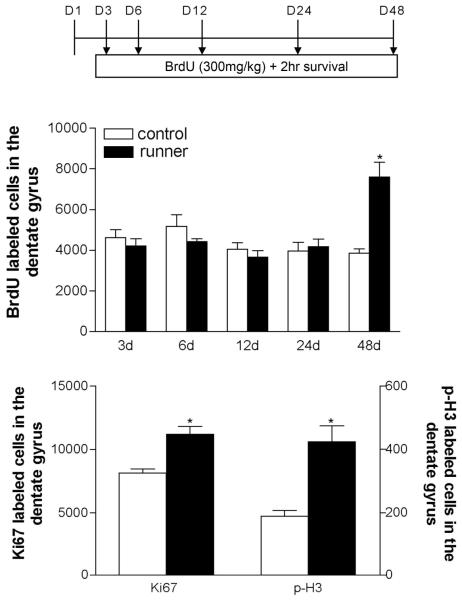
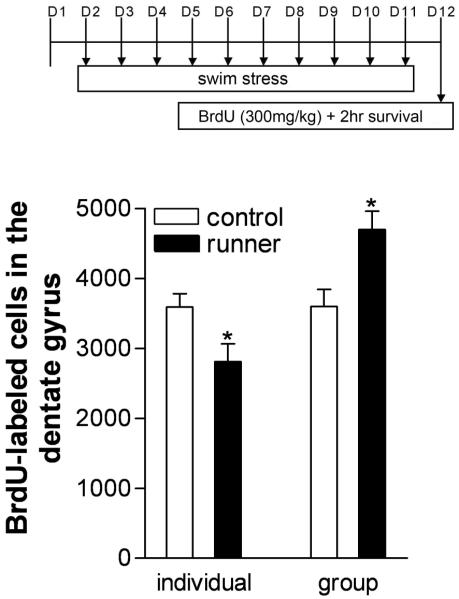
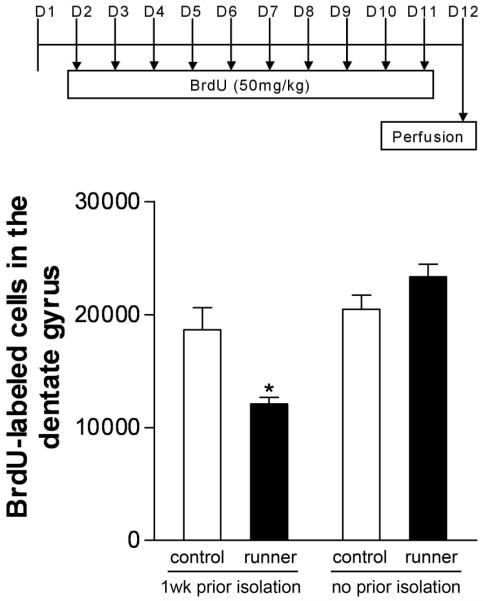
Social isolation can exacerbate the negative consequences of stress and increase the risk of developing psychopathology. However, the influence of living alone on experiences generally considered to be beneficial to the brain, such as physical exercise, remains unknown. We report here that individual housing precludes the positive influence of short-term running on adult neurogenesis in the hippocampus of rats and, in the presence of additional stress, suppresses the generation of new neurons. Individual housing also influenced corticosterone levels--runners in both housing conditions had elevated corticosterone during the active phase, but individually housed runners had higher levels of this hormone in response to stress. Moreover, lowering corticosterone levels converted the influence of short-term running on neurogenesis in individually housed rats from negative to positive. These results suggest that, in the absence of social interaction, a normally beneficial experience can exert a potentially deleterious influence on the brain.
Figures
References
-
- Selye H. The stress of life. McGraw-Hill; New York: 1976.
-
- Roy MP, Steptoe A, Kirschbaum C. Life events and social support as moderators of individual differences in cardiovascular and cortisol reactivity. J Pers Soc Psychol. 1998;75:1273–1281. - PubMed
-
- Weiss JM. Effects of coping behavior with and without a feedback signal on stress pathology in rats. J Comp Physiol Psychol. 1971;77:22–30. - PubMed
-
- Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci. 2005;8:365–371. - PubMed
-
- Weiss IC, Pryce CR, Jongen-Relo AL, Nanz-Bahr NI, Feldon J. Effect of social isolation on stress-related behavioral and neuroendocrine state in the rat. Beh Brain Res. 2004;152:279–295. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
