

Roles for HLA and KIR polymorphisms in natural killer cell repertoire selection and modulation of effector function
- PMID: 16533882
- PMCID: PMC2118260
- DOI: 10.1084/jem.20051884
Roles for HLA and KIR polymorphisms in natural killer cell repertoire selection and modulation of effector function
Erratum in
- J Exp Med. 2006 Apr 17;203(4):1131
Abstract
Interactions between killer cell immunoglobulin-like receptors (KIRs) and human leukocyte antigen (HLA) class I ligands regulate the development and response of human natural killer (NK) cells. Natural selection drove an allele-level group A KIR haplotype and the HLA-C1 ligand to unusually high frequency in the Japanese, who provide a particularly informative population for investigating the mechanisms by which KIR and HLA polymorphism influence NK cell repertoire and function. HLA class I ligands increase the frequencies of NK cells expressing cognate KIR, an effect modified by gene dose, KIR polymorphism, and the presence of other cognate ligand-receptor pairs. The five common Japanese KIR3DLI allotypes have distinguishable inhibitory capacity, frequency of cellular expression, and level of cell surface expression as measured by antibody binding. Although KIR haplotypes encoding 3DL1*001 or 3DL1*005, the strongest inhibitors, have no activating KIR, the dominant haplotype encodes a moderate inhibitor, 3DL1*01502, plus functional forms of the activating receptors 2DL4 and 2DS4. In the population, certain combinations of KIR and HLA class I ligand are overrepresented or underrepresented in women, but not men, and thus influence female fitness and survival. These findings show how KIR-HLA interactions shape the genetic and phenotypic KIR repertoires for both individual humans and the population.
Figures
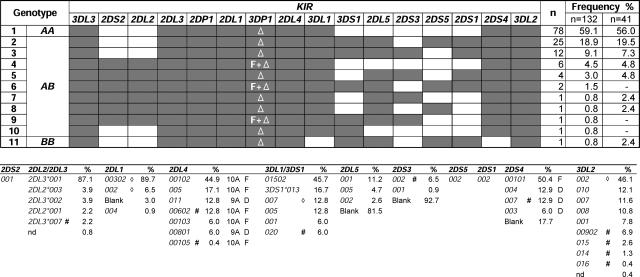
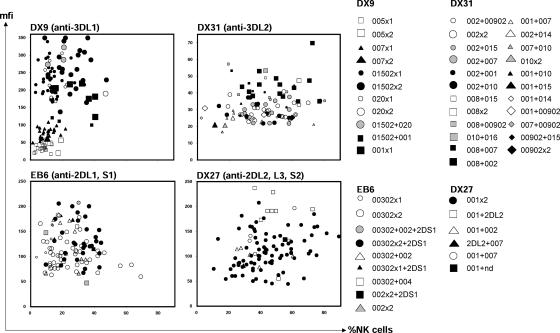
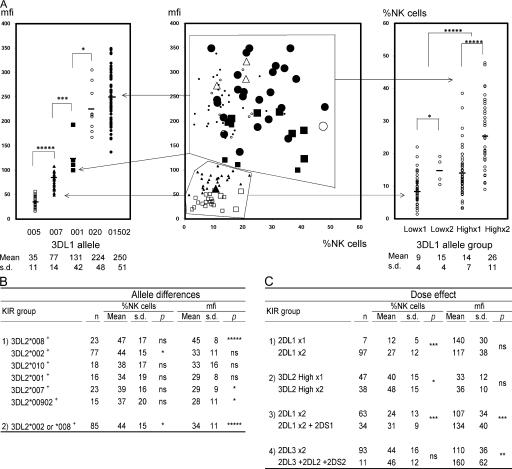
 : *001+*01502, Δ: *01502+*020, □: *005 × 1,
: *001+*01502, Δ: *01502+*020, □: *005 × 1,  : *005 × 2, ▴: *007 × 1,
: *005 × 2, ▴: *007 × 1,  : *007 × 2, •: *01502 × 1,
: *007 × 2, •: *01502 × 1,  : *01502 × 2, ◯: *020 × 1,
: *01502 × 2, ◯: *020 × 1,  : *020 × 2. The staining as a result of the “low-binding allotypes” (3DL1*005 and *007) is distinguished from that of the “high-binding allotypes” (3DL1*001, *01502, and *020) by the gates. The left panel is a one-dimensional plot of mfi showing how each of the five 3DL1 allotypes corresponds to a different range and mean value of DX9 binding. The right panel is a one-dimensional plot of percentage of NK cells binding DX9 in which the panel members were divided into four groups according to 3DL1 genotype: one low-binding allotype, two low-binding allotypes, one high-binding allotype, and two high-binding allotypes. The frequency of NK cells expressing 3DL1 increases with gene dose. (B) Correlations between flow cytometric analysis (%NK cells and mfi) with DX31 and the different 3DL2 allotypes. Pairwise comparisons were made between phenotypes with and without each 3DL2 allotype in “1),” or with and without the allotype group comprising 3DL2*002 and *008 in “2).” (C) The effects of gene dose on %NK cells and mfi observed with EB6, DX31, and DX27. Pairwise comparisons were made in phenotypes between the two donor groups that differed in gene content as indicated from “1)” to “4).” One copy of a gene is shown by x1, two copies are shown by x2. *, P < 0.05; **, P < 0.01; ***, P < 0.005; and *****, P < 0.0005.
: *020 × 2. The staining as a result of the “low-binding allotypes” (3DL1*005 and *007) is distinguished from that of the “high-binding allotypes” (3DL1*001, *01502, and *020) by the gates. The left panel is a one-dimensional plot of mfi showing how each of the five 3DL1 allotypes corresponds to a different range and mean value of DX9 binding. The right panel is a one-dimensional plot of percentage of NK cells binding DX9 in which the panel members were divided into four groups according to 3DL1 genotype: one low-binding allotype, two low-binding allotypes, one high-binding allotype, and two high-binding allotypes. The frequency of NK cells expressing 3DL1 increases with gene dose. (B) Correlations between flow cytometric analysis (%NK cells and mfi) with DX31 and the different 3DL2 allotypes. Pairwise comparisons were made between phenotypes with and without each 3DL2 allotype in “1),” or with and without the allotype group comprising 3DL2*002 and *008 in “2).” (C) The effects of gene dose on %NK cells and mfi observed with EB6, DX31, and DX27. Pairwise comparisons were made in phenotypes between the two donor groups that differed in gene content as indicated from “1)” to “4).” One copy of a gene is shown by x1, two copies are shown by x2. *, P < 0.05; **, P < 0.01; ***, P < 0.005; and *****, P < 0.0005.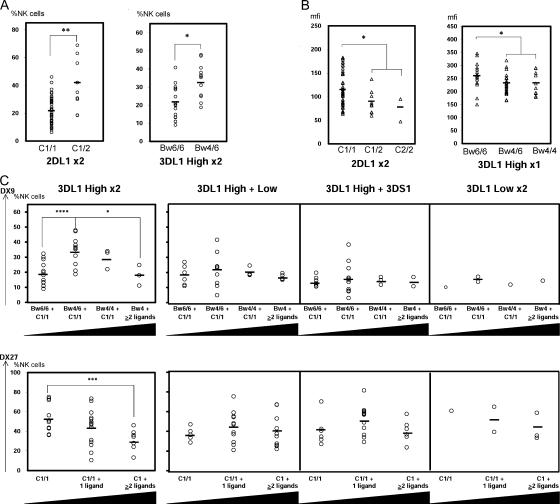
References
-
- Sun, P.D. 2003. Structure and function of natural-killer-cell receptors. Immunol. Res. 27:539–548. - PubMed
-
- Parham, P. 2005. MHC class I molecules and KIRs in human history, health and survival. Nat. Rev. Immunol. 5:201–214. - PubMed
-
- Tanamachi, D.M., D.C. Moniot, D. Cado, S.D. Liu, J.K. Hsia, and D.H. Raulet. 2004. Genomic Ly49A transgenes: basis of variegated Ly49A gene expression and identification of a critical regulatory element. J. Immunol. 172:1074–1082. - PubMed
-
- Held, W., B. Kunz, V. Ioannidis, and B. Lowin-Kropf. 1999. Mono-allelic Ly49 NK cell receptor expression. Semin. Immunol. 11:349–355. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
