

Ancient duplicated conserved noncoding elements in vertebrates: a genomic and functional analysis
- PMID: 16533910
- PMCID: PMC1457030
- DOI: 10.1101/gr.4143406
Ancient duplicated conserved noncoding elements in vertebrates: a genomic and functional analysis
Abstract
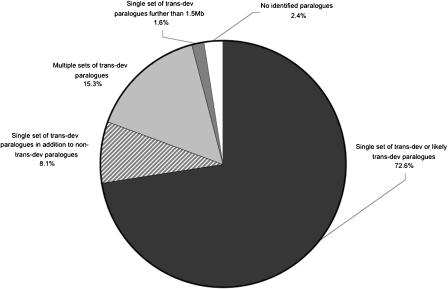
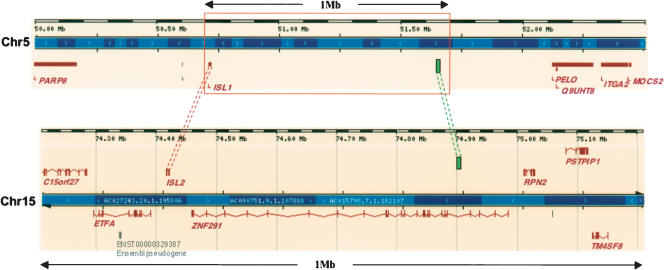
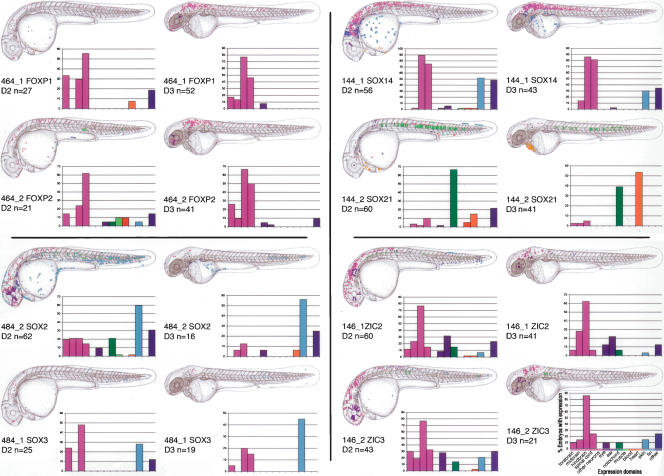
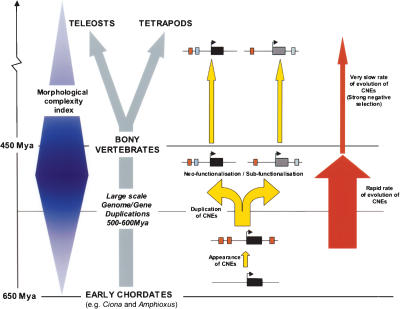
Fish-mammal genomic comparisons have proved powerful in identifying conserved noncoding elements likely to be cis-regulatory in nature, and the majority of those tested in vivo have been shown to act as tissue-specific enhancers associated with genes involved in transcriptional regulation of development. Although most of these elements share little sequence identity to each other, a small number are remarkably similar and appear to be the product of duplication events. Here, we searched for duplicated conserved noncoding elements in the human genome, using comparisons with Fugu to select putative cis-regulatory sequences. We identified 124 families of duplicated elements, each containing between two and five members, that are highly conserved within and between vertebrate genomes. In 74% of cases, we were able to assign a specific set of paralogous genes with annotation relating to transcriptional regulation and/or development to each family, thus removing much of the ambiguity in identifying associated genes. We find that duplicate elements have the potential to up-regulate reporter gene expression in a tissue-specific manner and that expression domains often overlap, but are not necessarily identical, between family members. Over two thirds of the families are conserved in duplicate in fish and appear to predate the large-scale duplication events thought to have occurred at the origin of vertebrates. We propose a model whereby gene duplication and the evolution of cis-regulatory elements can be considered in the context of increased morphological diversity and the emergence of the modern vertebrate body plan.
Figures




Similar articles
-
Comparative genomics using Fugu reveals insights into regulatory subfunctionalization.Genome Biol. 2007;8(4):R53. doi: 10.1186/gb-2007-8-4-r53. Genome Biol. 2007. PMID: 17428329 Free PMC article.
-
C-type lectin-like domains in Fugu rubripes.BMC Genomics. 2004 Aug 1;5(1):51. doi: 10.1186/1471-2164-5-51. BMC Genomics. 2004. PMID: 15285787 Free PMC article.
-
Comparative genomics using fugu: a tool for the identification of conserved vertebrate cis-regulatory elements.Bioessays. 2005 Jan;27(1):100-7. doi: 10.1002/bies.20134. Bioessays. 2005. PMID: 15612032 Review.
-
Deeply conserved chordate noncoding sequences preserve genome synteny but do not drive gene duplicate retention.Genome Res. 2009 Nov;19(11):2036-51. doi: 10.1101/gr.093237.109. Epub 2009 Aug 24. Genome Res. 2009. PMID: 19704032 Free PMC article.
-
[Identification and mapping of cis-regulatory elements within long genomic sequences].Mol Biol (Mosk). 2007 Sep-Oct;41(5):787-92. Mol Biol (Mosk). 2007. PMID: 18240562 Review. Russian.
Cited by
-
An ancient genomic regulatory block conserved across bilaterians and its dismantling in tetrapods by retrogene replacement.Genome Res. 2012 Apr;22(4):642-55. doi: 10.1101/gr.132233.111. Epub 2012 Jan 10. Genome Res. 2012. PMID: 22234889 Free PMC article.
-
Evaluation of IRX Genes and Conserved Noncoding Elements in a Region on 5p13.3 Linked to Families with Familial Idiopathic Scoliosis and Kyphosis.G3 (Bethesda). 2016 Jun 1;6(6):1707-12. doi: 10.1534/g3.116.029975. G3 (Bethesda). 2016. PMID: 27172222 Free PMC article.
-
Ancient Genomic Regulatory Blocks Are a Source for Regulatory Gene Deserts in Vertebrates after Whole-Genome Duplications.Mol Biol Evol. 2020 Oct 1;37(10):2857-2864. doi: 10.1093/molbev/msaa123. Mol Biol Evol. 2020. PMID: 32421818 Free PMC article.
-
Origin and evolution of spliceosomal introns.Biol Direct. 2012 Apr 16;7:11. doi: 10.1186/1745-6150-7-11. Biol Direct. 2012. PMID: 22507701 Free PMC article. Review.
-
Cis-regulatory landscapes in the evolution and development of the mammalian skull.Philos Trans R Soc Lond B Biol Sci. 2023 Jul 3;378(1880):20220079. doi: 10.1098/rstb.2022.0079. Epub 2023 May 15. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37183897 Free PMC article. Review.
References
-
- Aburomia R., Khaner O., Sidow A., Khaner O., Sidow A., Sidow A. Functional evolution in the ancestral lineage of vertebrates or when genomic complexity was wagging its morphological tail. J. Struct. Funct. Genomics. 2003;3:45–52. - PubMed
-
- Altschul S.F., Madden T.L., Schaffer A.A., Zhang J., Zhang Z., Miller W., Lipman D.J., Madden T.L., Schaffer A.A., Zhang J., Zhang Z., Miller W., Lipman D.J., Schaffer A.A., Zhang J., Zhang Z., Miller W., Lipman D.J., Zhang J., Zhang Z., Miller W., Lipman D.J., Zhang Z., Miller W., Lipman D.J., Miller W., Lipman D.J., Lipman D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–3402. - PMC - PubMed
-
- Aparicio S., Chapman J., Stupka E., Putnam N., Chia J.M., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., Chapman J., Stupka E., Putnam N., Chia J.M., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., Stupka E., Putnam N., Chia J.M., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., Putnam N., Chia J.M., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., Chia J.M., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., Dehal P., Christoffels A., Rash S., Hoon S., Smit A., Christoffels A., Rash S., Hoon S., Smit A., Rash S., Hoon S., Smit A., Hoon S., Smit A., Smit A., et al. Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science. 2002;297:1301–1310. - PubMed
-
- Arnone M.I., Davidson E.H., Davidson E.H. The hardwiring of development: Organization and function of genomic regulatory systems. Development. 1997;124:1851–1864. - PubMed
-
- Barton L.M., Gottgens B., Gering M., Gilbert J.G., Grafham D., Rogers J., Bentley D., Patient R., Green A.R., Gottgens B., Gering M., Gilbert J.G., Grafham D., Rogers J., Bentley D., Patient R., Green A.R., Gering M., Gilbert J.G., Grafham D., Rogers J., Bentley D., Patient R., Green A.R., Gilbert J.G., Grafham D., Rogers J., Bentley D., Patient R., Green A.R., Grafham D., Rogers J., Bentley D., Patient R., Green A.R., Rogers J., Bentley D., Patient R., Green A.R., Bentley D., Patient R., Green A.R., Patient R., Green A.R., Green A.R. Regulation of the stem cell leukemia (SCL) gene: A tale of two fishes. Proc. Natl. Acad. Sci. 2001;98:6747–6752. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources