

The avian coronavirus infectious bronchitis virus undergoes direct low-pH-dependent fusion activation during entry into host cells
- PMID: 16537586
- PMCID: PMC1440383
- DOI: 10.1128/JVI.80.7.3180-3188.2006
The avian coronavirus infectious bronchitis virus undergoes direct low-pH-dependent fusion activation during entry into host cells
Abstract
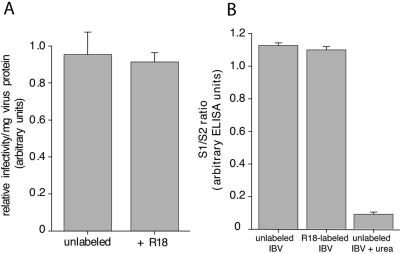
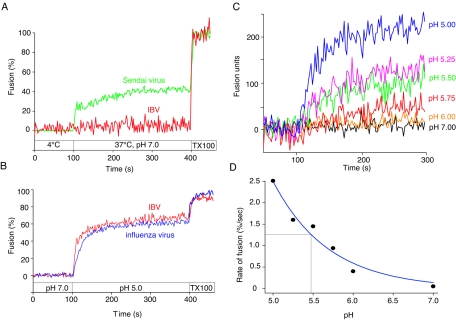
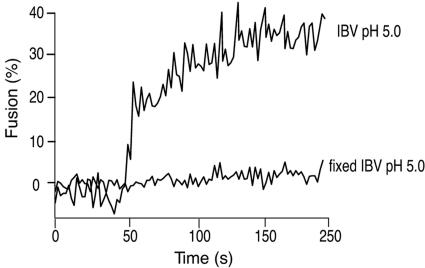
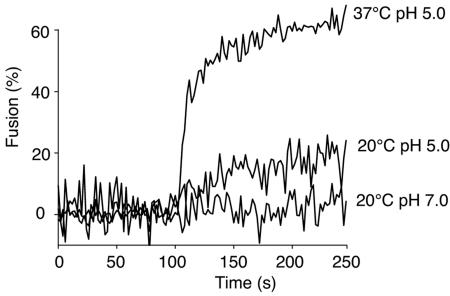
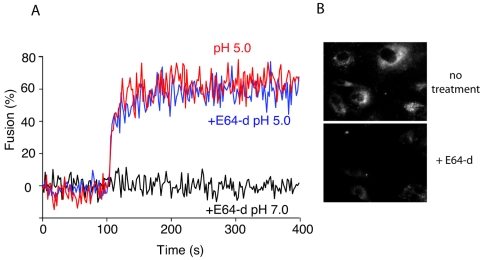
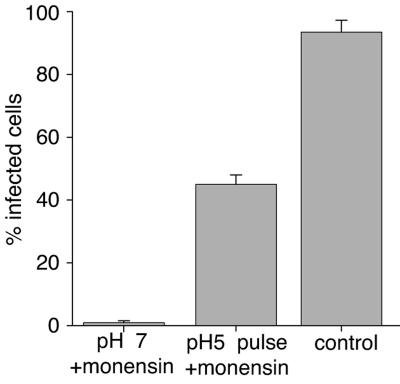
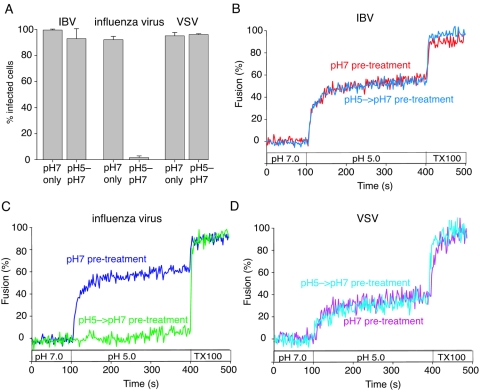
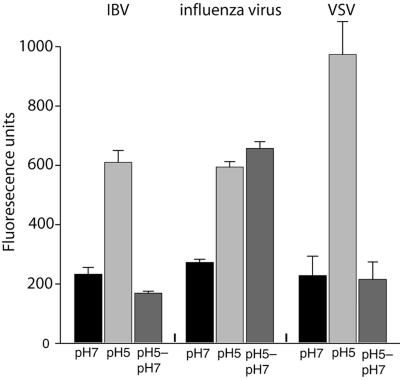
Coronaviruses are the causative agents of respiratory disease in humans and animals, including severe acute respiratory syndrome. Fusion of coronaviruses is generally thought to occur at neutral pH, although there is also evidence for a role of acidic endosomes during entry of a variety of coronaviruses. Therefore, the molecular basis of coronavirus fusion during entry into host cells remains incompletely defined. Here, we examined coronavirus-cell fusion and entry employing the avian coronavirus infectious bronchitis virus (IBV). Virus entry into cells was inhibited by acidotropic bases and by other inhibitors of pH-dependent endocytosis. We carried out fluorescence-dequenching fusion assays of R18-labeled virions and show that for IBV, coronavirus-cell fusion occurs in a low-pH-dependent manner, with a half-maximal rate of fusion occurring at pH 5.5. Fusion was reduced, but still occurred, at lower temperatures (20 degrees C). We observed no effect of inhibitors of endosomal proteases on the fusion event. These data are the first direct measure of virus-cell fusion for any coronavirus and demonstrate that the coronavirus IBV employs a direct, low-pH-dependent virus-cell fusion activation reaction. We further show that IBV was not inactivated, and fusion was unaffected, by prior exposure to pH 5.0 buffer. Virions also showed evidence of reversible conformational changes in their surface proteins, indicating that aspects of the fusion reaction may be reversible in nature.
Figures

References
-
- Beaudette, F. R., and C. R. Hudson. 1937. Cultivation of the virus of infectious bronchitis. J. Am. Vet. Med. Assoc. 90:51-60.
-
- Bosch, B. J., B. E. Martina, R. Van Der Zee, J. Lepault, B. J. Haijema, C. Versluis, A. J. Heck, R. De Groot, A. D. Osterhaus, and P. J. Rottier. 2004. Severe acute respiratory syndrome coronavirus (SARS-CoV) infection inhibition using spike protein heptad repeat-derived peptides. Proc. Natl. Acad. Sci. USA 101:8455-8460. - PMC - PubMed
-
- Cavanagh, D. 2005. Coronaviridae: a review of coronaviruses and toroviruses, p. 1-54. In A. Schmidt, M. H. Wolff, and O. Weber (ed.), Coronaviruses with special emphasis on first insights concerning SARS. Birkhauser Verlag, Basel, Switzerland.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
