

UL26-deficient human cytomegalovirus produces virions with hypophosphorylated pp28 tegument protein that is unstable within newly infected cells
- PMID: 16537622
- PMCID: PMC1440364
- DOI: 10.1128/JVI.80.7.3541-3548.2006
UL26-deficient human cytomegalovirus produces virions with hypophosphorylated pp28 tegument protein that is unstable within newly infected cells
Abstract
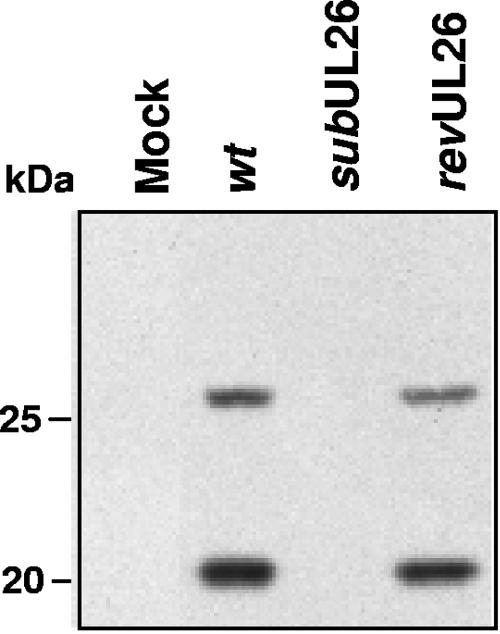
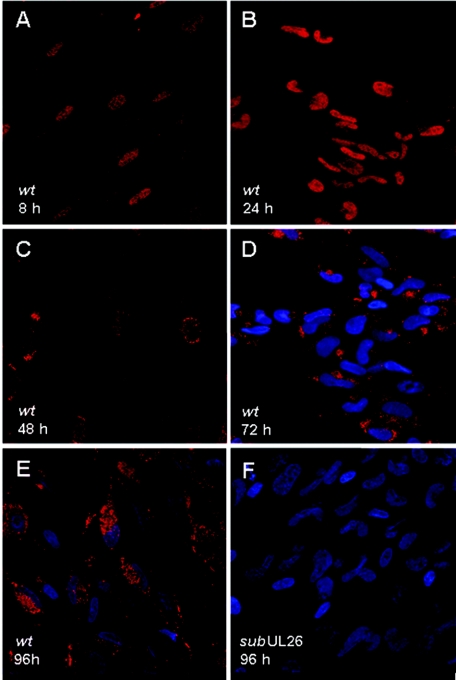
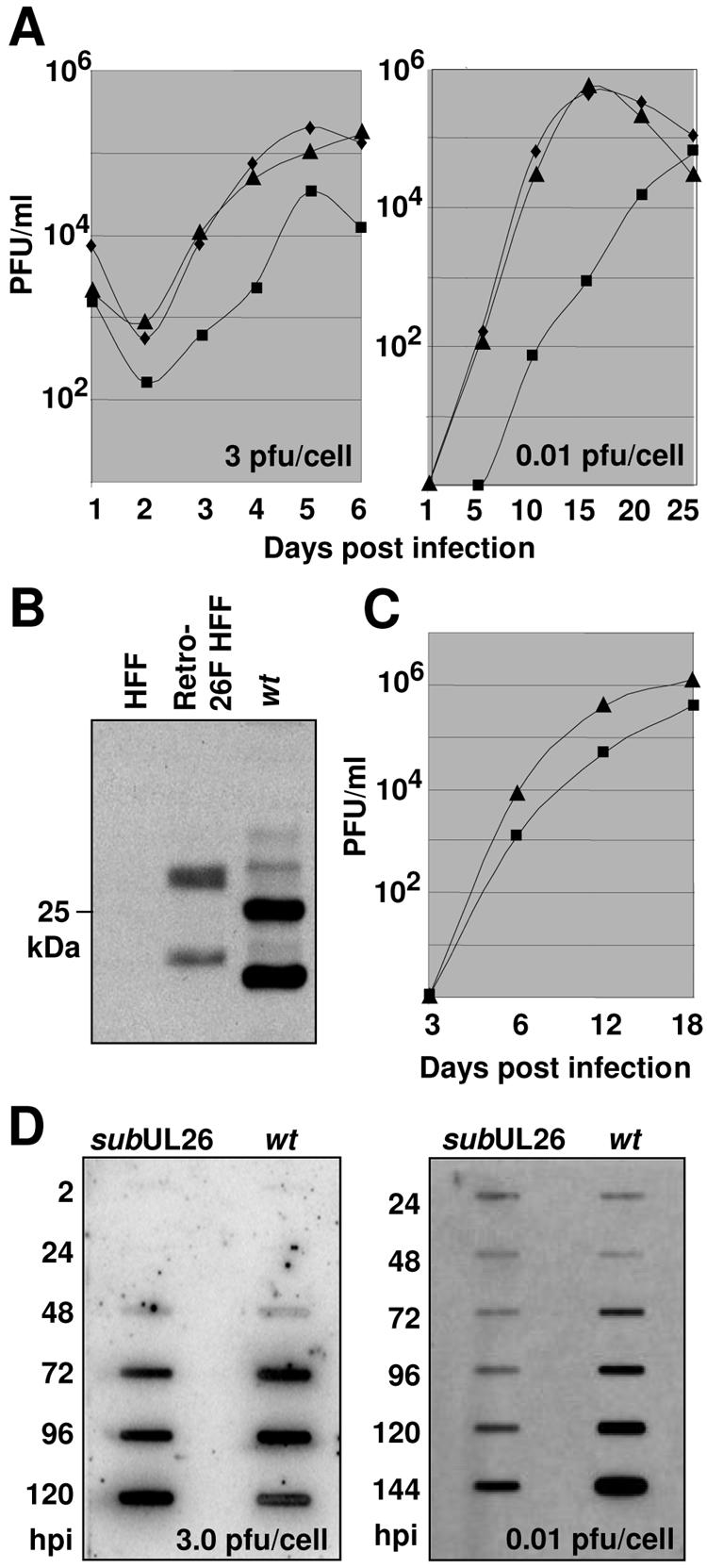
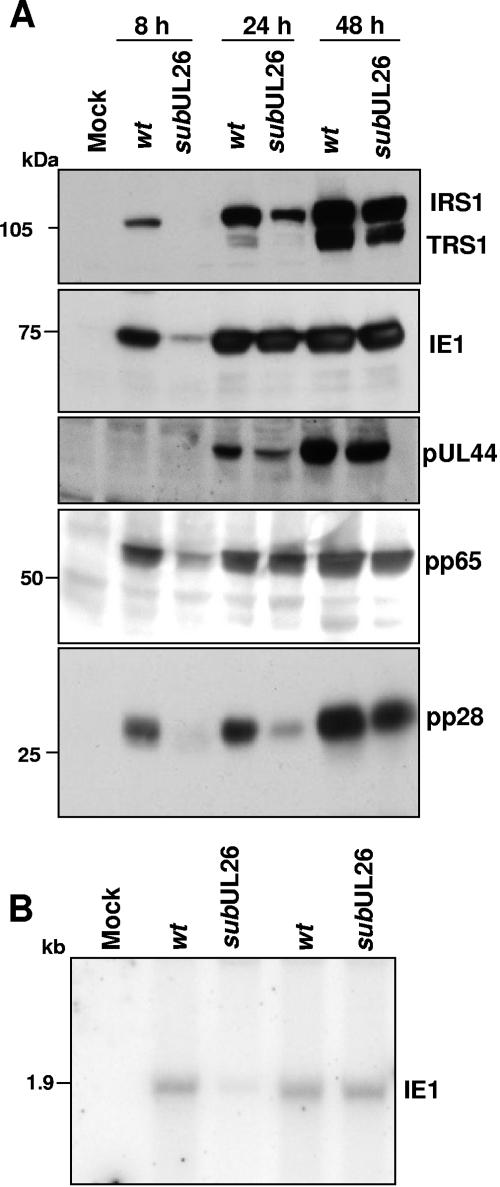
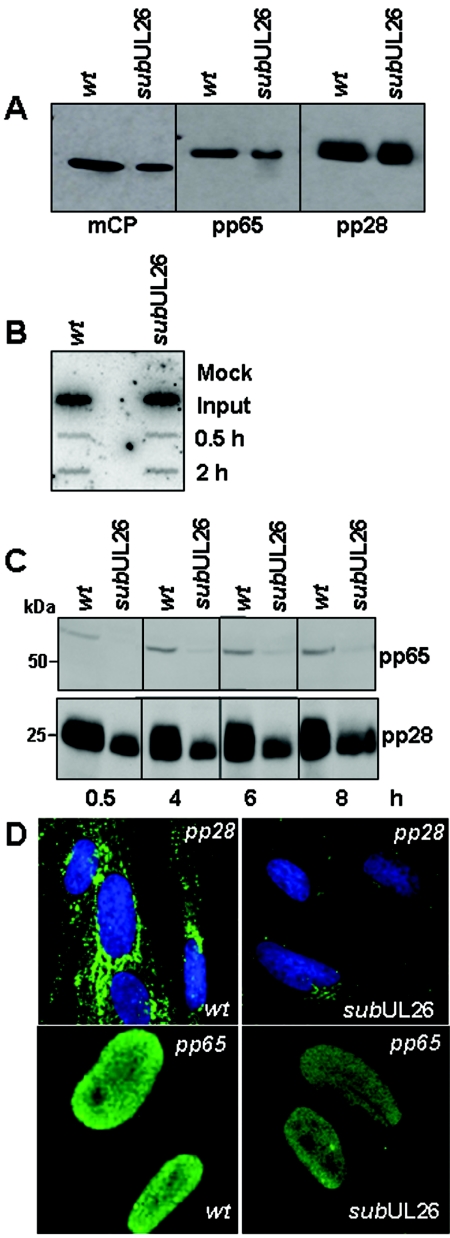
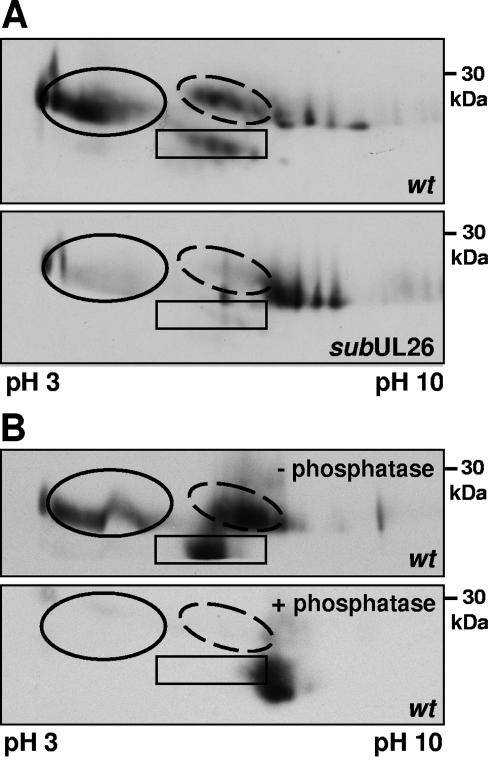
The human cytomegalovirus UL26 open reading frame encodes proteins of 21 and 27 kDa that result from the use of two different in-frame initiation codons. The UL26 protein is a constituent of the virion and thus is delivered to cells upon viral entry. We have characterized a mutant of human cytomegalovirus in which the UL26 open reading frame has been deleted. The UL26 deletion mutant has a profound growth defect, the magnitude of which is dependent on the multiplicity of infection. Two very early defects were discovered. First, even though they were present in normal amounts within mutant virions, the UL99-coded pp28 and UL83-coded pp65 tegument proteins were present in reduced amounts at the earliest times assayed within newly infected cells; second, there was a delay in immediate-early mRNA and protein accumulation. Further analysis revealed that although wild-type levels of the pp28 tegument protein were present in UL26 deletion mutant virions, the protein was hypophosphorylated. We conclude that the UL26 protein influences the normal phosphorylation of at least pp28 in virions and possibly additional tegument proteins. We propose that the hypophosphorylation of tegument proteins causes their destabilization within newly infected cells, perhaps disrupting the normal detegumentation process and leading to a delay in the onset of immediate-early gene expression.
Figures
References
-
- Anderson, M. L. M. 1999. Nucleic acid hybridization. Springer, New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
