

Cotranscriptional recognition of human intronic box H/ACA snoRNAs occurs in a splicing-independent manner
- PMID: 16537900
- PMCID: PMC1430331
- DOI: 10.1128/MCB.26.7.2540-2549.2006
Cotranscriptional recognition of human intronic box H/ACA snoRNAs occurs in a splicing-independent manner
Abstract
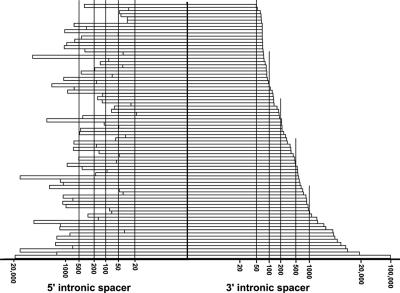
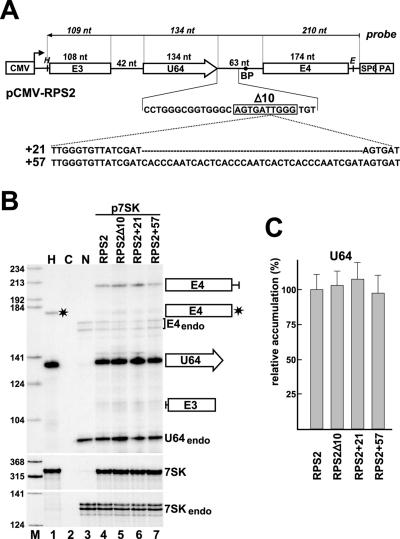
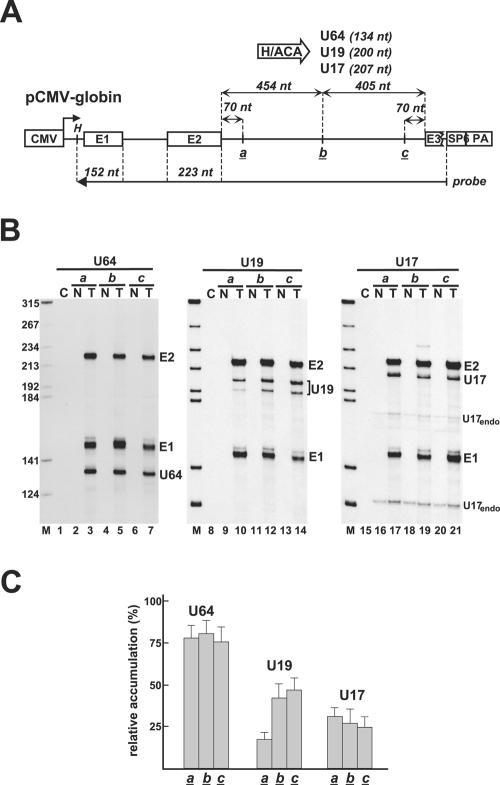
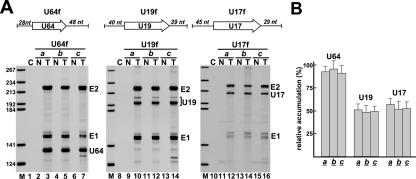
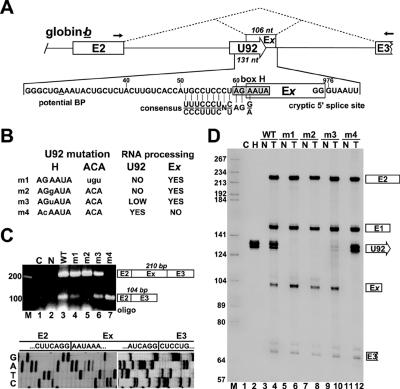
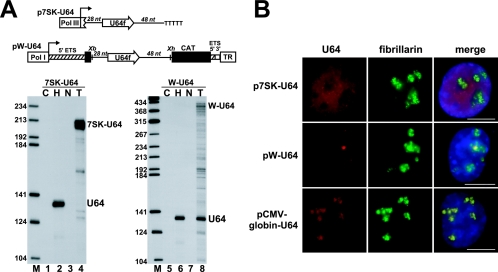
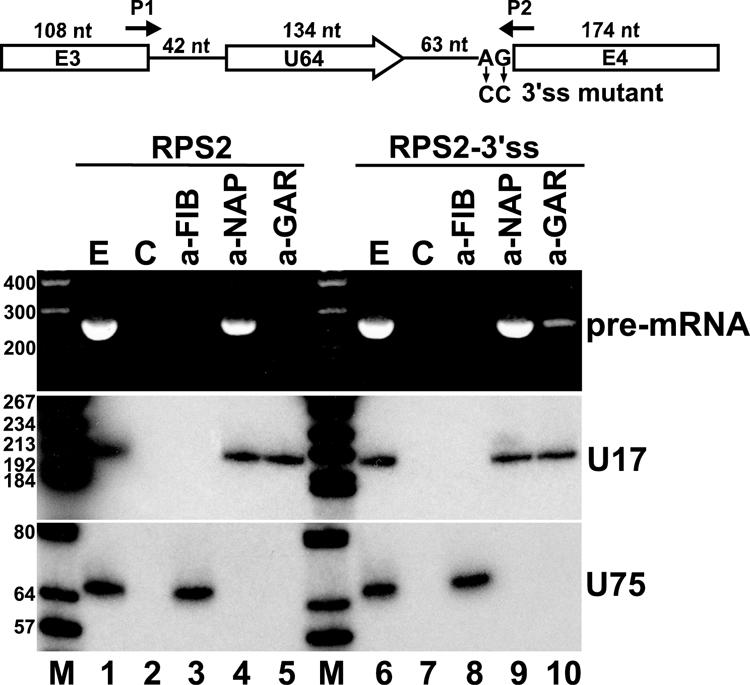
Processing from pre-mRNA introns is a widespread mechanism to generate human box C/D and H/ACA snoRNAs. Recent studies revealed that an optimal position relative to the 3' splice site is important for efficient processing of most box C/D snoRNAs and that assembly of box C/D snoRNPs is stimulated by splicing factors likely bound to the branch point region. Here we have investigated the processing of another major class of human intron-encoded RNAs, the box H/ACA snoRNAs. Analysis of 80 H/ACA RNA genes revealed that human H/ACA RNAs possess no preferential localization close to the 3' or 5' splice site. In vivo processing experiments confirmed that H/ACA intronic snoRNAs are processed in a position-independent manner, indicating that there is no synergy between H/ACA RNA processing and splicing. We also showed that recognition of intronic H/ACA snoRNAs and assembly of pre-snoRNPs is an early event that occurs during transcription elongation parallel with pre-mRNA splice site selection. Finally, we found that efficient processing and correct nucleolar localization of the human U64 H/ACA snoRNA requires RNA polymerase II-mediated synthesis of the U64 precursor. This suggests that polymerase II-associated factors direct the efficient assembly and determine the correct subnuclear trafficking of human H/ACA snoRNPs.
Figures
Similar articles
-
The position of yeast snoRNA-coding regions within host introns is essential for their biosynthesis and for efficient splicing of the host pre-mRNA.RNA. 2007 Jan;13(1):138-50. doi: 10.1261/rna.251907. Epub 2006 Nov 29. RNA. 2007. PMID: 17135484 Free PMC article.
-
Naf1 p is a box H/ACA snoRNP assembly factor.RNA. 2002 Dec;8(12):1502-14. RNA. 2002. PMID: 12515383 Free PMC article.
-
A combined computational and experimental analysis of two families of snoRNA genes from Caenorhabditis elegans, revealing the expression and evolution pattern of snoRNAs in nematodes.Genomics. 2007 Apr;89(4):490-501. doi: 10.1016/j.ygeno.2006.12.002. Epub 2007 Jan 11. Genomics. 2007. PMID: 17222528
-
SnoRNP biogenesis meets Pre-mRNA splicing.Mol Cell. 2006 Sep 15;23(6):775-6. doi: 10.1016/j.molcel.2006.08.023. Mol Cell. 2006. PMID: 16973429 Review.
-
The vertebrate E1/U17 small nucleolar ribonucleoprotein particle.J Cell Biochem. 2006 Jun 1;98(3):486-95. doi: 10.1002/jcb.20821. J Cell Biochem. 2006. PMID: 16475166 Review.
Cited by
-
Assembly and trafficking of box C/D and H/ACA snoRNPs.RNA Biol. 2017 Jun 3;14(6):680-692. doi: 10.1080/15476286.2016.1243646. Epub 2016 Oct 7. RNA Biol. 2017. PMID: 27715451 Free PMC article. Review.
-
Integrating snoRNP assembly with mRNA biogenesis.EMBO Rep. 2006 Jun;7(6):590-2. doi: 10.1038/sj.embor.7400715. EMBO Rep. 2006. PMID: 16741502 Free PMC article. No abstract available.
-
Dysregulation of Small Nucleolar RNAs in B-Cell Malignancies.Biomedicines. 2022 May 24;10(6):1229. doi: 10.3390/biomedicines10061229. Biomedicines. 2022. PMID: 35740251 Free PMC article. Review.
-
Bcd1p controls RNA loading of the core protein Nop58 during C/D box snoRNP biogenesis.RNA. 2019 Apr;25(4):496-506. doi: 10.1261/rna.067967.118. Epub 2019 Jan 30. RNA. 2019. PMID: 30700579 Free PMC article.
-
Novel pathological variants of NHP2 affect N-terminal domain flexibility, protein stability, H/ACA Ribonucleoprotein (RNP) complex formation and telomerase activity.Hum Mol Genet. 2023 Sep 16;32(19):2901-2912. doi: 10.1093/hmg/ddad114. Hum Mol Genet. 2023. PMID: 37440454 Free PMC article.
References
-
- Ambros, V. 2004. The functions of animal microRNAs. Nature 431:350-355. - PubMed
-
- Bachellerie, J. P., J. Cavaillé, and A. Hüttenhofer. 2002. The expanding snoRNA world. Biochimie 84:775-790. - PubMed
-
- Bartel, D. P. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116:281-297. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources