

Role for centromeric heterochromatin and PML nuclear bodies in the cellular response to foreign DNA
- PMID: 16537904
- PMCID: PMC1430340
- DOI: 10.1128/MCB.26.7.2583-2594.2006
Role for centromeric heterochromatin and PML nuclear bodies in the cellular response to foreign DNA
Erratum in
- Mol Cell Biol. 2006 May;26(10):3997
Abstract
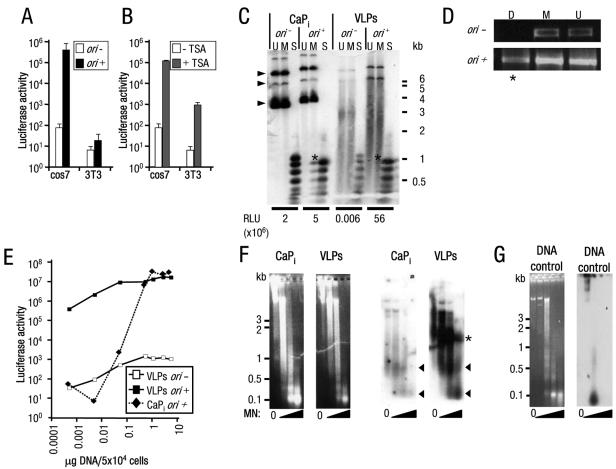
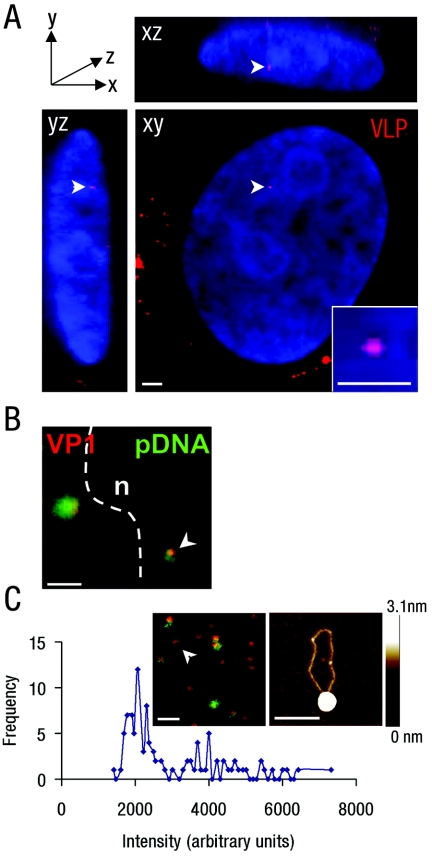
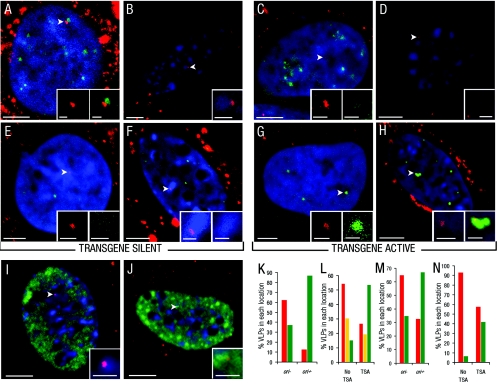
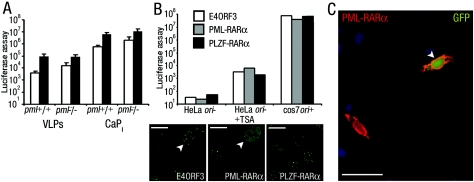
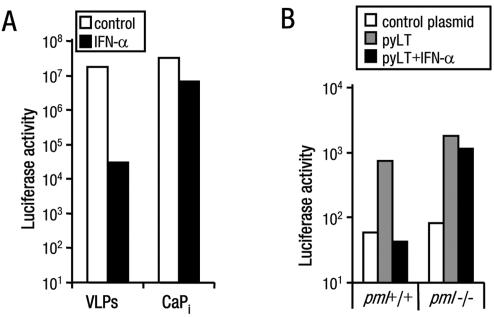
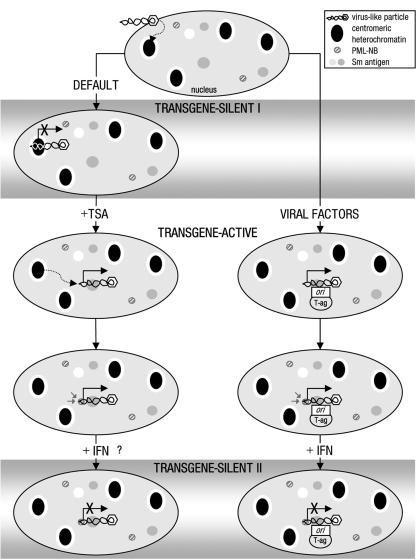
Nuclear spatial positioning plays an important role in the epigenetic regulation of eukaryotic gene expression. Here we show a role for nuclear spatial positioning in regulating episomal transgenes that are delivered by virus-like particles (VLPs). VLPs mediate the delivery of plasmid DNA (pDNA) to cell nuclei but lack viral factors involved in initiating and regulating transcription. By tracking single fluorescently labeled VLPs, coupled with luciferase reporter gene assays, we found that VLPs transported pDNA to cell nuclei efficiently but transgenes were immediately silenced by the cell. An investigation of the nuclear location of fluorescent VLPs revealed that the pDNAs were positioned next to centromeric heterochromatin. The activation of transcription by providing viral factors or inhibiting histone deacetylase activity resulted in the localization to euchromatin regions. Further, the activation of transcription induced the recruitment of PML nuclear bodies (PML-NBs) to the VLPs. This association did not play a role in regulating transgene expression, but PML protein was necessary for the inhibition of transgene expression with alpha interferon (IFN-alpha). These results support a model whereby cells can prevent foreign gene expression at two levels: by positioning transgenes next to centromeric heterochromatin or, if that is overcome, via the type I IFN response facilitated by PML-NB recruitment.
Figures
Similar articles
-
Transcriptional regulation is affected by subnuclear targeting of reporter plasmids to PML nuclear bodies.Mol Cell Biol. 2006 Dec;26(23):8814-25. doi: 10.1128/MCB.00636-06. Epub 2006 Sep 11. Mol Cell Biol. 2006. PMID: 16966371 Free PMC article.
-
Cell cycle behavior of human HP1 subtypes: distinct molecular domains of HP1 are required for their centromeric localization during interphase and metaphase.J Cell Sci. 2003 Aug 15;116(Pt 16):3327-38. doi: 10.1242/jcs.00635. Epub 2003 Jul 2. J Cell Sci. 2003. PMID: 12840071
-
The efficiency of nuclear plasmid DNA delivery is a critical determinant of transgene expression at the single cell level.J Gene Med. 2010 Jan;12(1):77-85. doi: 10.1002/jgm.1406. J Gene Med. 2010. PMID: 19890943
-
The vagaries of variegating transgenes.Bioessays. 1996 Nov;18(11):919-23. doi: 10.1002/bies.950181111. Bioessays. 1996. PMID: 8939070 Review.
-
Molecular Complexes at Euchromatin, Heterochromatin and Centromeric Chromatin.Int J Mol Sci. 2021 Jun 28;22(13):6922. doi: 10.3390/ijms22136922. Int J Mol Sci. 2021. PMID: 34203193 Free PMC article. Review.
Cited by
-
The histone deacetylase inhibitor Entinostat enhances polymer-mediated transgene expression in cancer cell lines.Biotechnol Bioeng. 2016 Jun;113(6):1345-1356. doi: 10.1002/bit.25898. Epub 2015 Dec 30. Biotechnol Bioeng. 2016. PMID: 26614912 Free PMC article.
-
Recognition and suppression of transfected plasmids by protein ZNF511-PRAP1, a potential molecular barrier to transgene expression.Mol Ther. 2011 Aug;19(8):1478-86. doi: 10.1038/mt.2011.80. Epub 2011 May 3. Mol Ther. 2011. PMID: 21540836 Free PMC article.
-
Alterations of Nuclear Architecture and Epigenetic Signatures during African Swine Fever Virus Infection.Viruses. 2015 Sep 15;7(9):4978-96. doi: 10.3390/v7092858. Viruses. 2015. PMID: 26389938 Free PMC article.
-
An Adenovirus DNA Replication Factor, but Not Incoming Genome Complexes, Targets PML Nuclear Bodies.J Virol. 2015 Nov 25;90(3):1657-67. doi: 10.1128/JVI.02545-15. Print 2016 Feb 1. J Virol. 2015. PMID: 26608315 Free PMC article.
-
Imaging single virus particles on the surface of cell membranes by high-resolution scanning surface confocal microscopy.Biophys J. 2008 May 15;94(10):4089-94. doi: 10.1529/biophysj.107.112524. Epub 2008 Jan 16. Biophys J. 2008. PMID: 18199668 Free PMC article.
References
-
- An, K., A. Q. Paulsen, M. B. Tilley, and R. A. Consigli. 2000. Use of electron microscopic and immunogold labeling techniques to determine polyomavirus recombinant VP1 capsid-like particles entry into mouse 3T6 cell nucleus. J. Virol. Methods 90:91-97. - PubMed
-
- Baxter, J., M. Merkenschlager, and A. G. Fisher. 2002. Nuclear organisation and gene expression. Curr. Opin. Cell Biol. 14:372-376. - PubMed
-
- Belmont, A. S., S. Dietzel, A. C. Nye, Y. G. Strukov, and T Tumbar. 1999. Large-scale chromatin structure and function. Curr. Opin. Cell Biol. 11:307-311. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous