

PRISM/PRDM6, a transcriptional repressor that promotes the proliferative gene program in smooth muscle cells
- PMID: 16537907
- PMCID: PMC1430312
- DOI: 10.1128/MCB.26.7.2626-2636.2006
PRISM/PRDM6, a transcriptional repressor that promotes the proliferative gene program in smooth muscle cells
Abstract
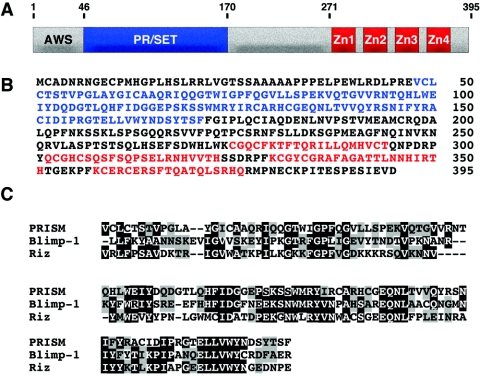
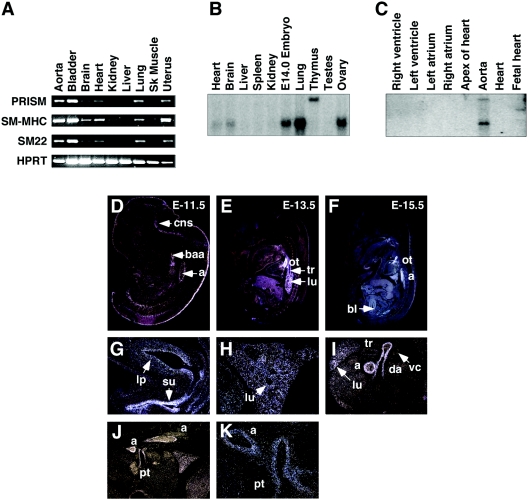
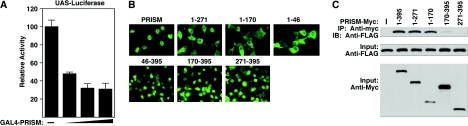
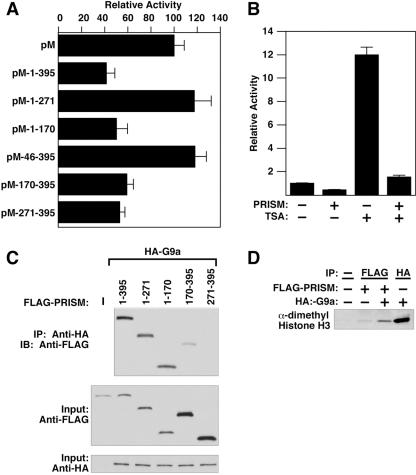
Smooth muscle cells (SMCs) display remarkable phenotypic diversity and plasticity and can readily switch between proliferative and differentiated states in response to extracellular cues. In an effort to identify novel transcriptional regulators of smooth muscle phenotypes, we compared the gene expression profiles of arterial and venous SMCs by microarray-based transcriptional profiling. Among numerous genes displaying distinct expression patterns in these two SMC types, we discovered an expressed sequence tag encoding a previously uncharacterized zinc finger protein belonging to the PRDM (PRDI-BF1 and RIZ homology domain) family of chromatin-remodeling proteins and named it PRISM (PR domain in smooth muscle). PRISM is expressed in a variety of smooth muscle-containing tissues and displays especially robust expression in the cardiac outflow tract and descending aorta during embryogenesis. PRISM is localized to the nucleus and contains an amino-terminal PR domain and four Krüppel-like zinc fingers at the carboxy terminus. We show that PRISM acts as a transcriptional repressor by interacting with class I histone deacetylases and the G9a histone methyltransferase, thereby identifying PRISM as a novel SMC-restricted epigenetic regulator. Overexpression of PRISM in cultured primary SMCs induces genes associated with the proliferative smooth muscle phenotype while repressing regulators of differentiation, including myocardin and GATA-6. Conversely, small interfering RNA-mediated knockdown of PRISM slows cell growth and induces myocardin, GATA-6, and markers of SMC differentiation. We conclude that PRISM acts as a novel epigenetic regulator of SMC phenotypic plasticity by suppressing differentiation and maintaining the proliferative potential of vascular SMCs.
Figures


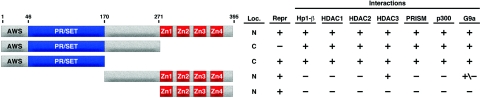
Similar articles
-
PRDM6 is enriched in vascular precursors during development and inhibits endothelial cell proliferation, survival, and differentiation.J Mol Cell Cardiol. 2008 Jan;44(1):47-58. doi: 10.1016/j.yjmcc.2007.06.008. Epub 2007 Jun 30. J Mol Cell Cardiol. 2008. PMID: 17662997 Free PMC article.
-
Kruppel-like factor 4, Elk-1, and histone deacetylases cooperatively suppress smooth muscle cell differentiation markers in response to oxidized phospholipids.Am J Physiol Cell Physiol. 2008 Nov;295(5):C1175-82. doi: 10.1152/ajpcell.00288.2008. Epub 2008 Sep 3. Am J Physiol Cell Physiol. 2008. PMID: 18768922 Free PMC article.
-
Pitx2 is functionally important in the early stages of vascular smooth muscle cell differentiation.J Cell Biol. 2008 May 5;181(3):461-73. doi: 10.1083/jcb.200711145. J Cell Biol. 2008. PMID: 18458156 Free PMC article.
-
Vascular smooth muscle cell phenotypic plasticity: focus on chromatin remodelling.Cardiovasc Res. 2012 Jul 15;95(2):147-55. doi: 10.1093/cvr/cvs098. Epub 2012 Feb 22. Cardiovasc Res. 2012. PMID: 22362814 Free PMC article. Review.
-
Molecular control of vascular smooth muscle cell differentiation and phenotypic plasticity.Novartis Found Symp. 2007;283:174-91; discussion 191-3, 238-41. doi: 10.1002/9780470319413.ch14. Novartis Found Symp. 2007. PMID: 18300422 Review.
Cited by
-
Integrative single-cell analysis of cardiogenesis identifies developmental trajectories and non-coding mutations in congenital heart disease.Cell. 2022 Dec 22;185(26):4937-4953.e23. doi: 10.1016/j.cell.2022.11.028. Cell. 2022. PMID: 36563664 Free PMC article.
-
Two novel SNPs in the coding region of the bovine PRDM16 gene and its associations with growth traits.Mol Biol Rep. 2010 Jan;37(1):571-7. doi: 10.1007/s11033-009-9816-8. Mol Biol Rep. 2010. PMID: 19760096
-
Genome-wide analysis of sex-specific differences in the mother-child PELAGIE cohort exposed to organophosphate metabolites.Sci Rep. 2023 May 17;13(1):8003. doi: 10.1038/s41598-023-35113-8. Sci Rep. 2023. PMID: 37198424 Free PMC article.
-
PRDM6 promotes medulloblastoma by repressing chromatin accessibility and altering gene expression.Sci Rep. 2024 Jul 12;14(1):16074. doi: 10.1038/s41598-024-66811-6. Sci Rep. 2024. PMID: 38992221 Free PMC article.
-
A case study in cross-talk: the histone lysine methyltransferases G9a and GLP.Nucleic Acids Res. 2010 Jun;38(11):3503-11. doi: 10.1093/nar/gkq081. Epub 2010 Feb 16. Nucleic Acids Res. 2010. PMID: 20159995 Free PMC article. Review.
References
-
- Baxendale, S., C. Davison, C. Muxworthy, C. Wolff, P. W. Ingham, and S. Roy. 2004. The B-cell maturation factor Blimp-1 specifies vertebrate slow-twitch muscle fiber identity in response to Hedgehog signaling. Nat. Genet. 36:88-93. - PubMed
-
- Cremona, O., M. Muda, R. D. Appel, S. Frutiger, G. J. Hughes, D. F. Hochstrasser, A. Geinoz, and G. Gabbiani. 1995. Differential protein expression in aortic smooth muscle cells cultured from newborn and aged rats. Exp. Cell Res. 217:280-287. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials