

Zoledronic acid treatment impairs protein geranyl-geranylation for biological effects in prostatic cells
- PMID: 16539721
- PMCID: PMC1434759
- DOI: 10.1186/1471-2407-6-60
Zoledronic acid treatment impairs protein geranyl-geranylation for biological effects in prostatic cells
Abstract
Background: Nitrogen-containing bisphosphonates (N-BPs) have been designed to inhibit osteoclast-mediated bone resorption. However, it is now accepted that part of their anti-tumor activities is related to interference with the mevalonate pathway.
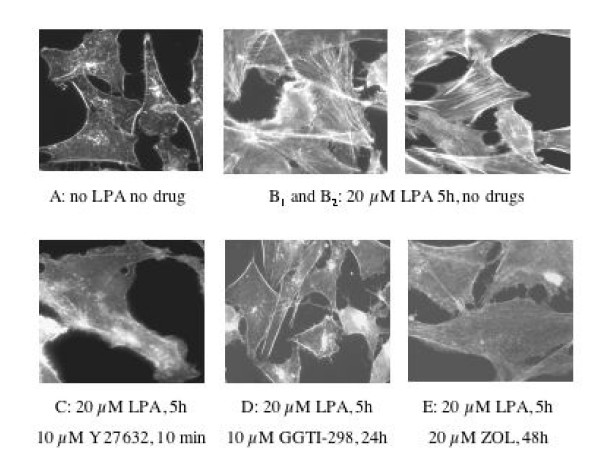
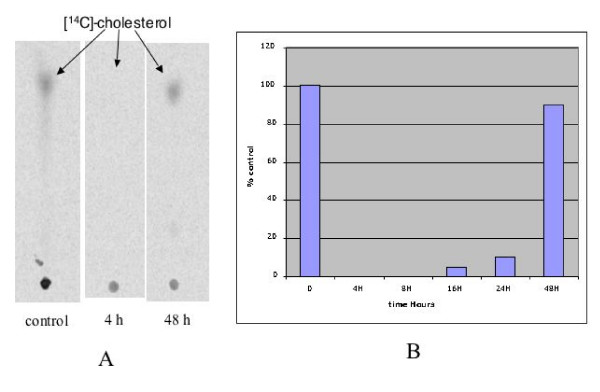
Methods: We investigated the effects of zoledronic acid (ZOL), on cell proliferation and protein isoprenylation in two tumoral (LnCAP, PC-3,), and one normal established (PNT1-A) prostatic cell line. To assess if inhibition of geranyl-geranylation by ZOL impairs the biological activity of RhoA GTPase, we studied the LPA-induced formation of stress fibers. The inhibitory effect of ZOL on geranyl geranyl transferase I was checked biochemically. Activity of ZOL on cholesterol biosynthesis was determined by measuring the incorporation of 14C mevalonate in cholesterol.
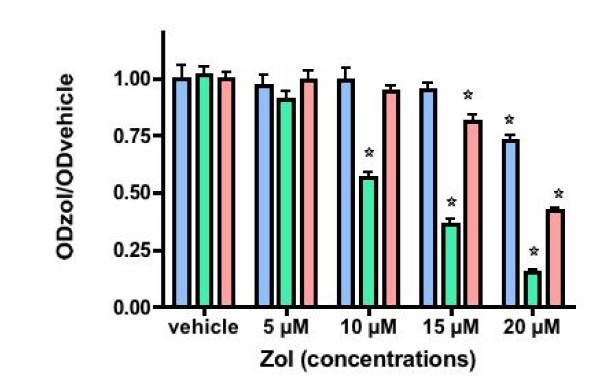
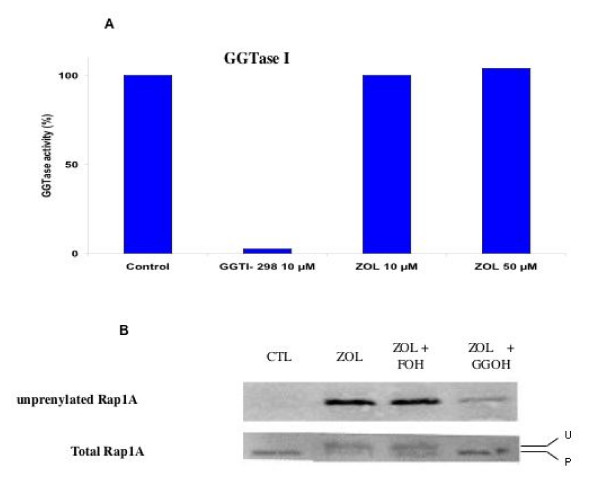
Results: ZOL induced dose-dependent inhibition of proliferation of all the three cell lines although it appeared more efficient on the untransformed PNT1A. Whatever the cell line, 20 microM ZOL-induced inhibition was reversed by geranyl-geraniol (GGOH) but neither by farnesol nor mevalonate. After 48 hours treatment of cells with 20 microM ZOL, geranyl-geranylation of Rap1A was abolished whereas farnesylation of HDJ-2 was unaffected. Inhibition of Rap1A geranyl-geranylation by ZOL was rescued by GGOH and not by FOH. Indeed, as observed with treatment by a geranyl-geranyl transferase inhibitor, treatment of PNT1-A cells with 20 microM ZOL prevented the LPA-induced formation of stress fibers. We checked that in vitro ZOL did not inhibit geranyl-geranyl-transferase I. ZOL strongly inhibited cholesterol biosynthesis up to 24 hours but at 48 hours 90% of this biosynthesis was rescued.
Conclusion: Although zoledronic acid is currently the most efficient bisphosphonate in metastatic prostate cancer management, its mechanism of action in prostatic cells remains unclear. We suggest in this work that although in first intention ZOL inhibits FPPsynthase its main biological actitivity is directed against protein Geranylgeranylation.
Figures
 PC3
PC3  PNT1A
PNT1A  LNCaP
LNCaP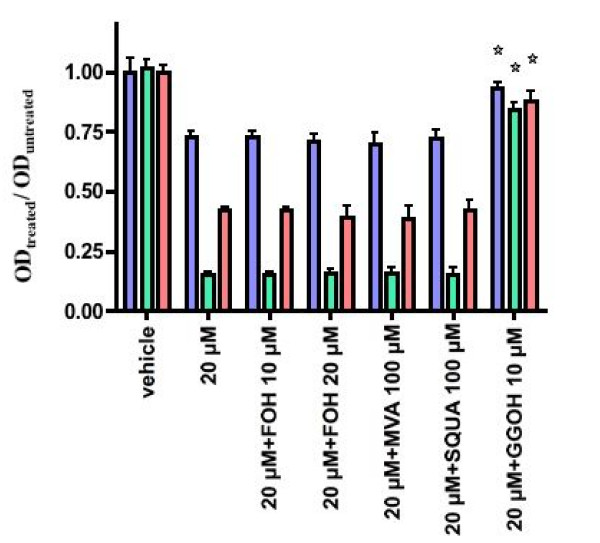 PC-3 PNT1A LNCaP
PC-3 PNT1A LNCaP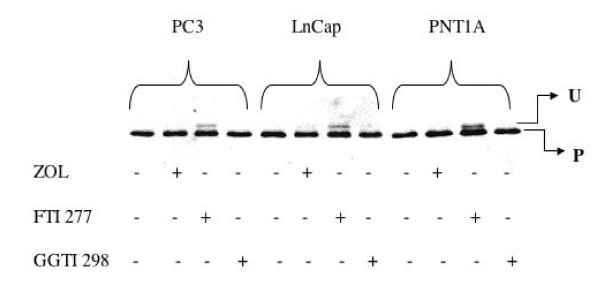
References
-
- Amin D, Cornell SA, Gustafson SK, Needle SJ, Ullrich JW, Bilder GE, Perrone MH. Bisphosphonates used for the treatment of bone disorders inhibit squalene synthase and cholesterol biosynthesis. J Lipid Res. 1992;33:1657–1663. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
