

Glial cells dilate and constrict blood vessels: a mechanism of neurovascular coupling
- PMID: 16540563
- PMCID: PMC2270788
- DOI: 10.1523/JNEUROSCI.4048-05.2006
Glial cells dilate and constrict blood vessels: a mechanism of neurovascular coupling
Abstract
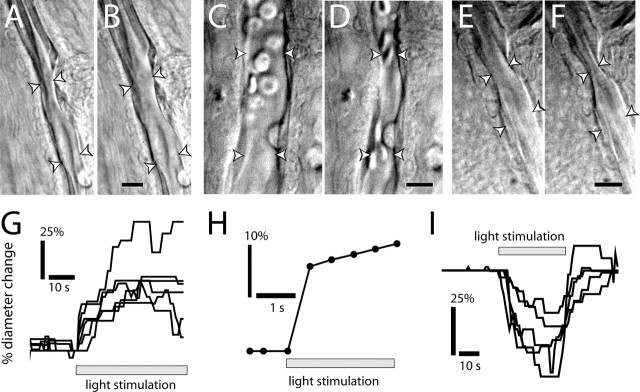
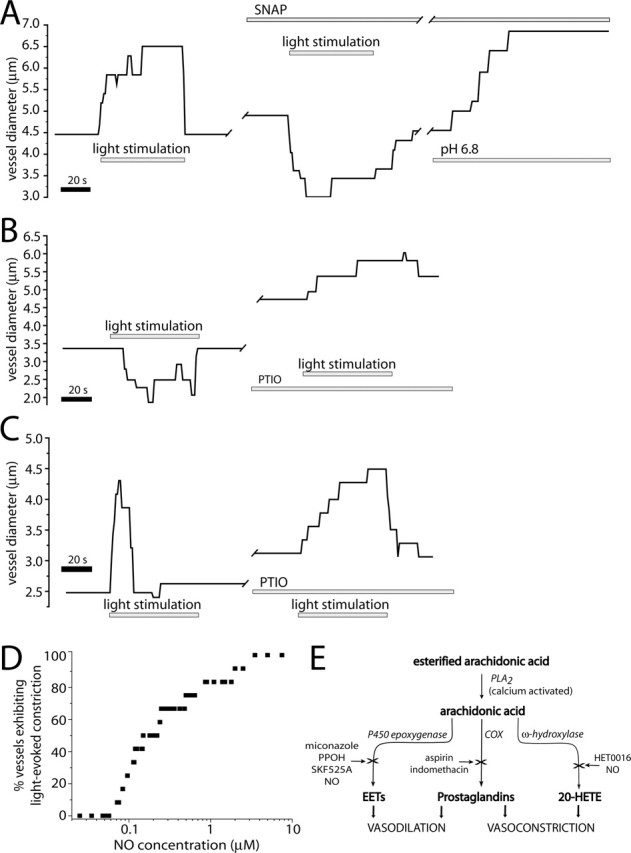
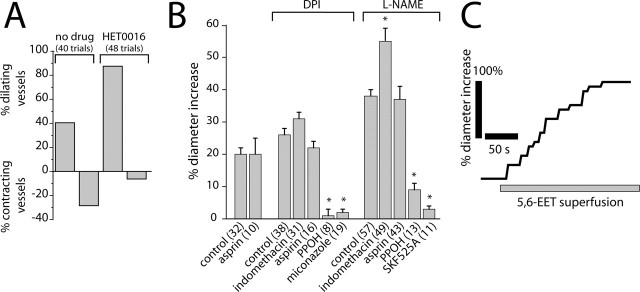
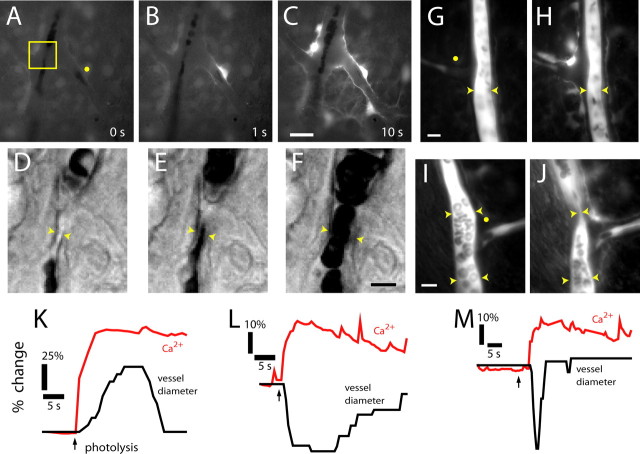
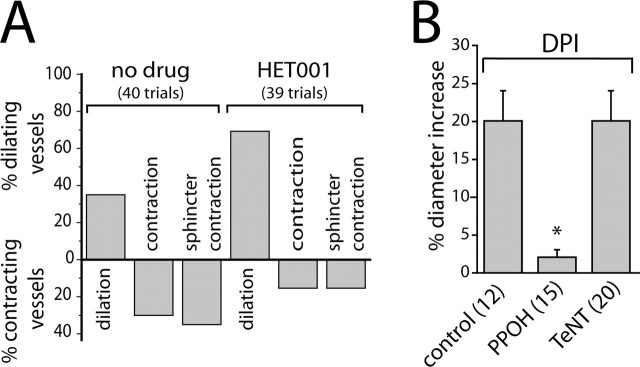
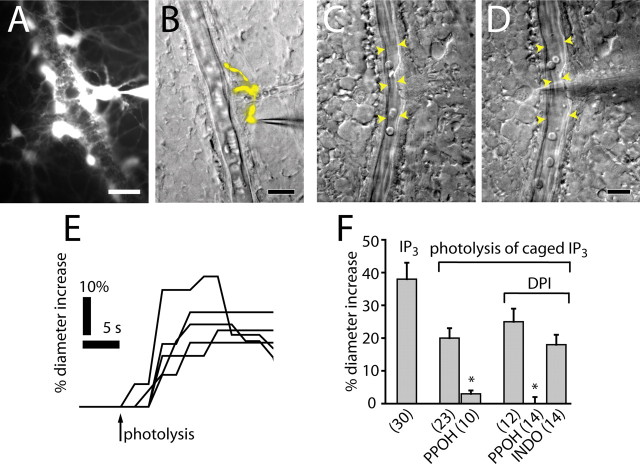
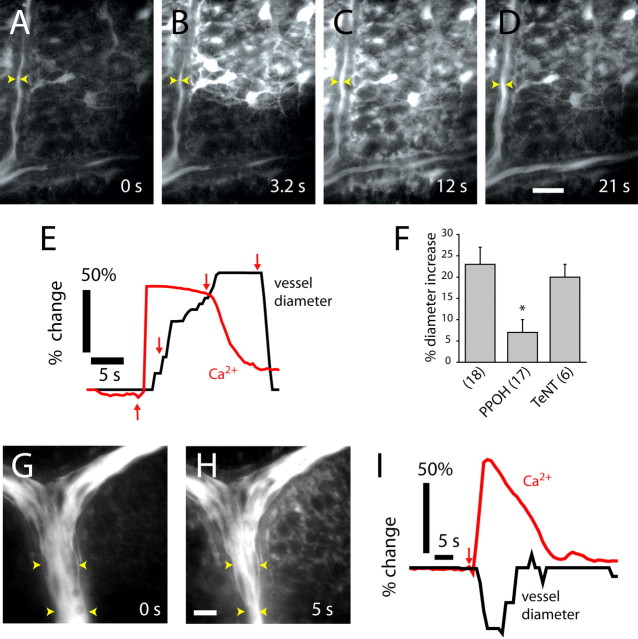
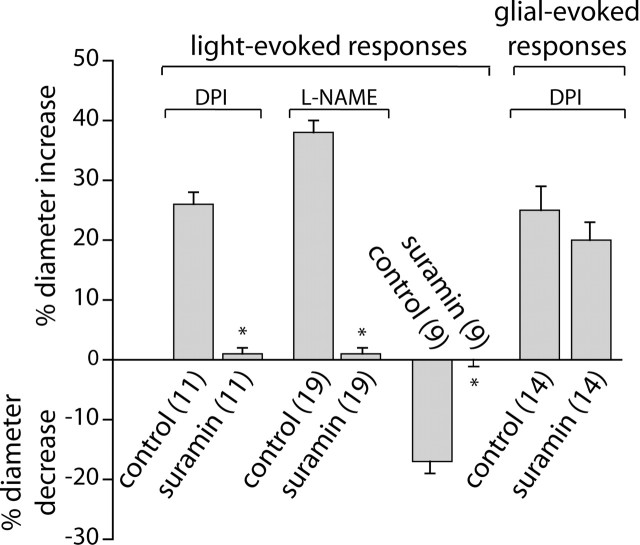
Neuronal activity evokes localized changes in blood flow. Although this response, termed neurovascular coupling, is widely used to monitor human brain function and diagnose pathology, the cellular mechanisms that mediate the response remain unclear. We investigated the contribution of glial cells to neurovascular coupling in the acutely isolated mammalian retina. We found that light stimulation and glial cell stimulation can both evoke dilation or constriction of arterioles. Light-evoked and glial-evoked vasodilations were blocked by inhibitors of cytochrome P450 epoxygenase, the synthetic enzyme for epoxyeicosatrienoic acids. Vasoconstrictions, in contrast, were blocked by an inhibitor of omega-hydroxylase, which synthesizes 20-hydroxyeicosatetraenoic acid. Nitric oxide influenced whether vasodilations or vasoconstrictions were produced in response to light and glial stimulation. Light-evoked vasoactivity was blocked when neuron-to-glia signaling was interrupted by a purinergic antagonist. These results indicate that glial cells contribute to neurovascular coupling and suggest that regulation of blood flow may involve both vasodilating and vasoconstricting components.
Figures
References
-
- Alkayed NJ, Narayanan J, Gebremedhin D, Medhora M, Roman RJ, Harder DR (1996). Molecular characterization of an arachidonic acid epoxygenase in rat brain astrocytes. Stroke 27:971–979. - PubMed
-
- Alonso-Galicia M, Sun CW, Falck JR, Harder DR, Roman RJ (1998). Contribution of 20-HETE to the vasodilator actions of nitric oxide in renal arteries. Am J Physiol Renal Physiol 275:F370–F378. - PubMed
-
- Alonso-Galicia M, Falck JR, Reddy KM, Roman RJ (1999). 20-HETE agonists and antagonists in the renal circulation. Am J Physiol Renal Physiol 277:F790–F796. - PubMed
-
- Amruthesh SC, Boerschel MF, McKinney JS, Willoughby KA, Ellis EF (1993). Metabolism of arachidonic acid to epoxyeicosatrienoic acids, hydroxyeicosatetraenoic acids, and prostaglandins in cultured rat hippocampal astrocytes. J Neurochem 61:150–159. - PubMed
-
- Anderson CM, Nedergaard M (2003). Astrocyte-mediated control of cerebral microcirculation. Trends Neurosci 26:340–344. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources