

Differential projections of excitatory and inhibitory dorsal horn interneurons relaying information from group II muscle afferents in the cat spinal cord
- PMID: 16540564
- PMCID: PMC1890019
- DOI: 10.1523/JNEUROSCI.5172-05.2006
Differential projections of excitatory and inhibitory dorsal horn interneurons relaying information from group II muscle afferents in the cat spinal cord
Abstract
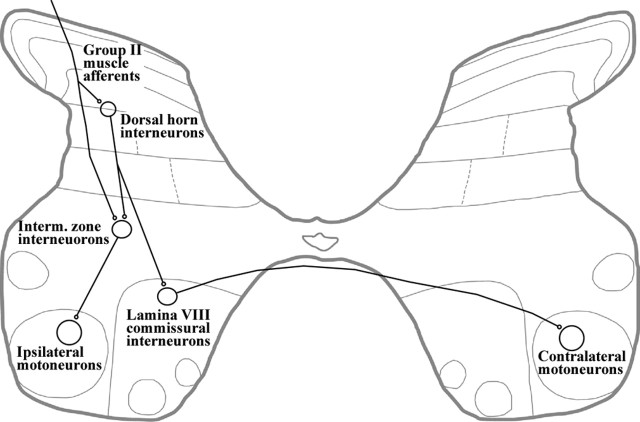
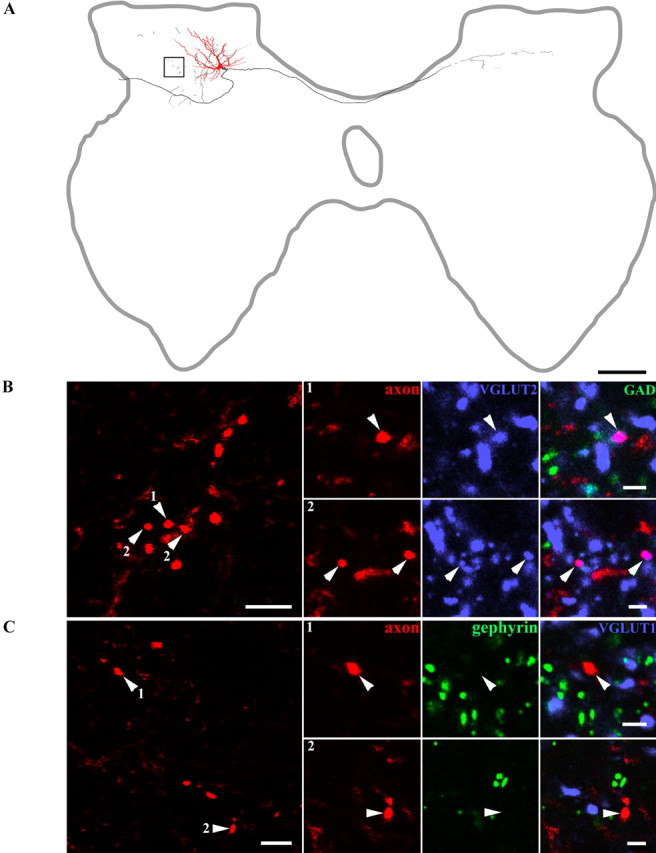
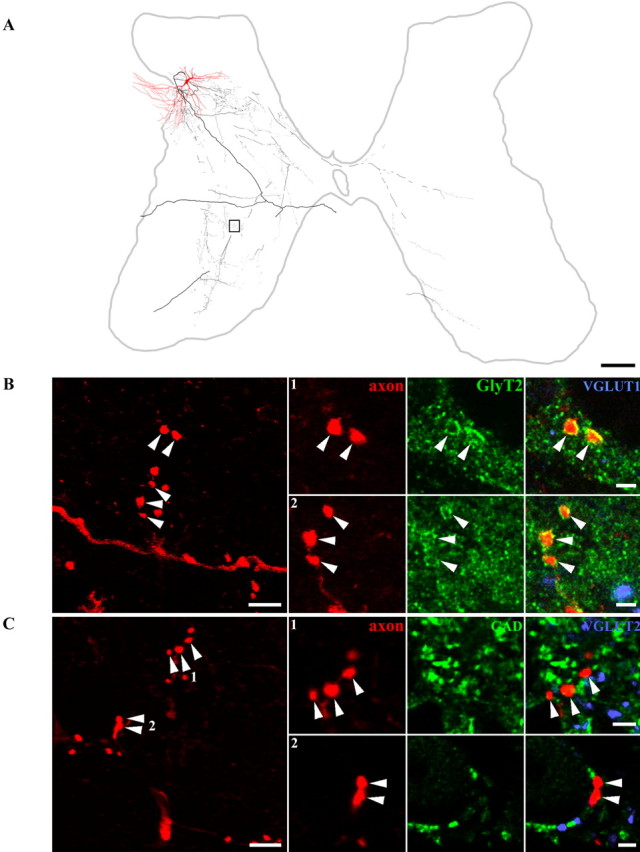
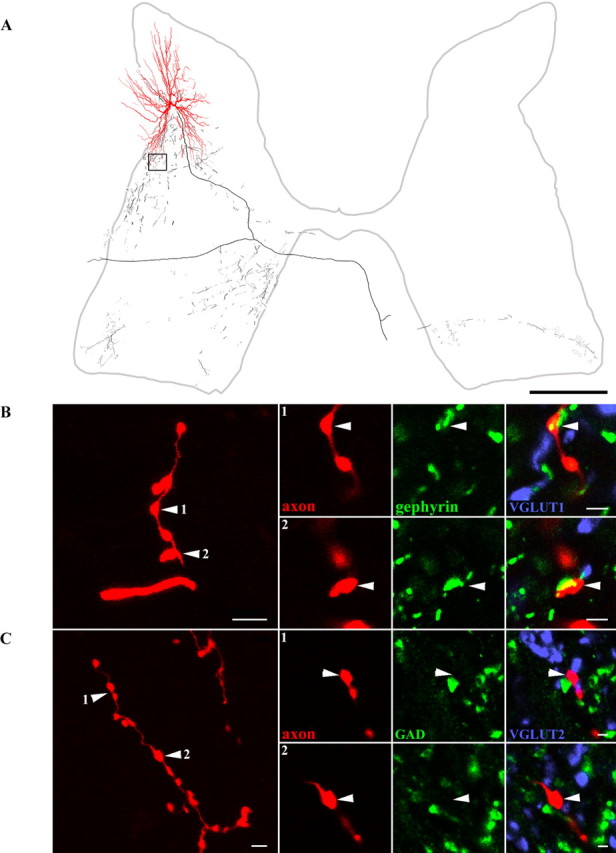
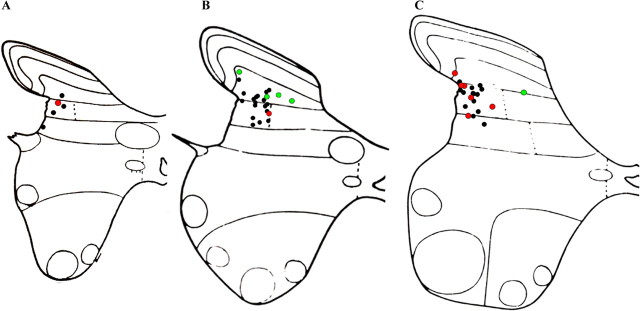
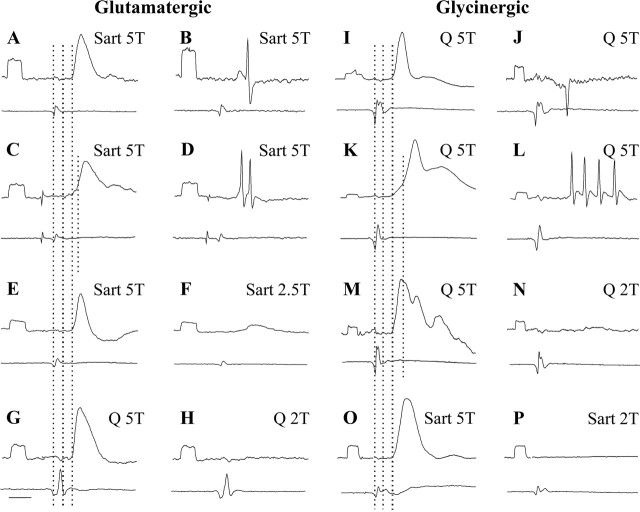
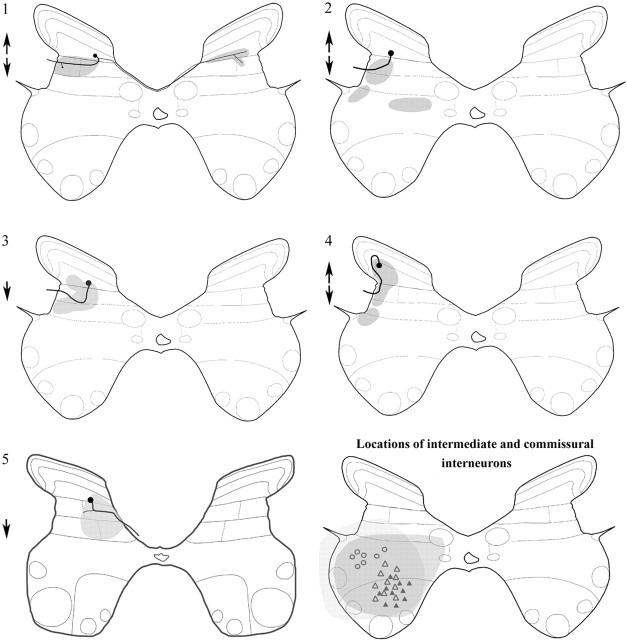
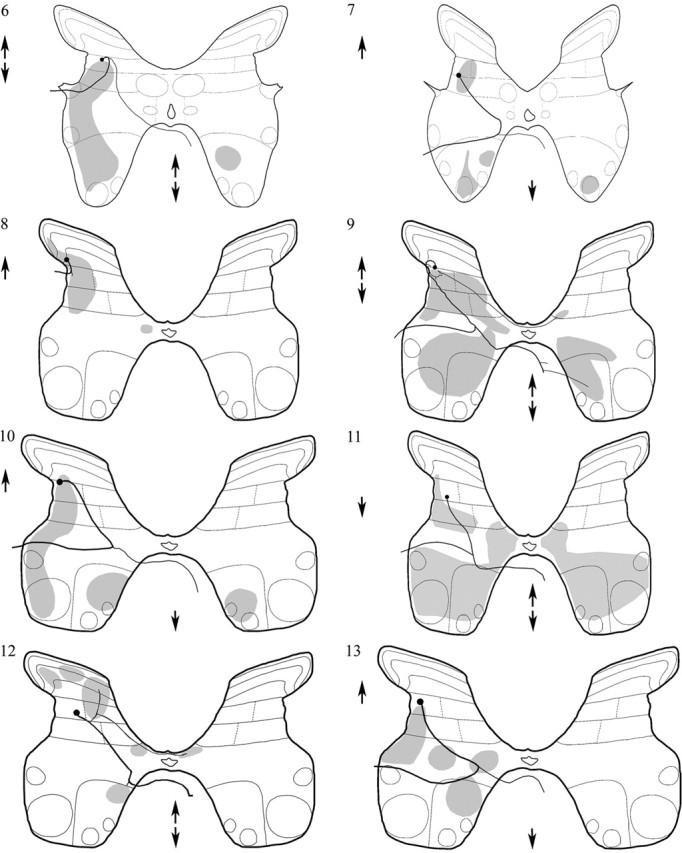
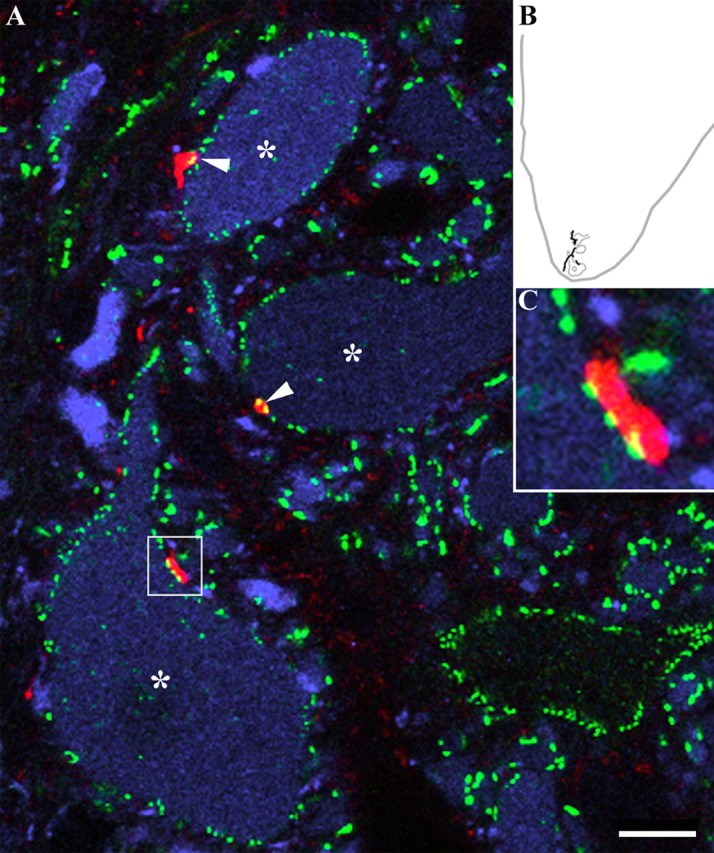
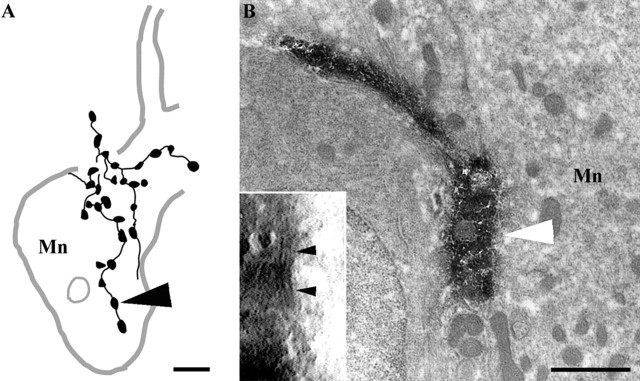
Dorsal horn interneurons with input from group II muscle spindle afferents are components of networks involved in motor control. Thirteen dorsal horn interneurons with monosynaptic group II input were characterized electrophysiologically and labeled intracellularly with Neurobiotin. Their axonal projections were traced, and neurotransmitter content was established by using immunocytochemistry. Two subpopulations were identified: five interneurons had axons that contained vesicular glutamate transporter 2 and hence were glutamatergic and excitatory. Terminals of the remaining eight interneurons were immunoreactive for the glycine transporter 2 or were apposed to gephyrin but did not contain the GABA-synthesizing enzyme glutamic acid decarboxylase and were therefore glycinergic and inhibitory. Excitatory cells were located mainly in the central region of lamina IV and had relatively small somata and restricted dendritic trees. In contrast, inhibitory interneurons were located more ventrally, in lamina V and had relatively larger somata and more extensive dendritic trees. Axonal projections of the two subpopulations differed considerably. Excitatory interneurons predominantly projected ipsilaterally, whereas most inhibitory interneurons projected both ipsilaterally and contralaterally. Three inhibitory axons formed contacts with large cholinergic cells in motor nuclei, thus revealing a novel direct coupling between inhibitory dorsal horn interneurons and motoneurons. The organization of the excitatory interneurons is consistent with current knowledge of reflex pathways to motoneurons, but the existence and connections of the inhibitory subpopulation could not be predicted from previous data. Our results indicate that these latter interneurons exercise widespread inhibitory control over a variety of cell types located on both sides of the spinal cord.
Figures
References
-
- Alvarez FJ, Dewey DE, Harrington DA, Fyffe RE (1997). Cell-type specific receptor organisation of glycine receptor clusters in the mammalian spinal cord. J Comp Neurol 397:150–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous