

Activation of the caspase 8 pathway mediates seizure-induced cell death in cultured hippocampal neurons
- PMID: 16542823
- PMCID: PMC1618926
- DOI: 10.1016/j.eplepsyres.2006.02.002
Activation of the caspase 8 pathway mediates seizure-induced cell death in cultured hippocampal neurons
Abstract
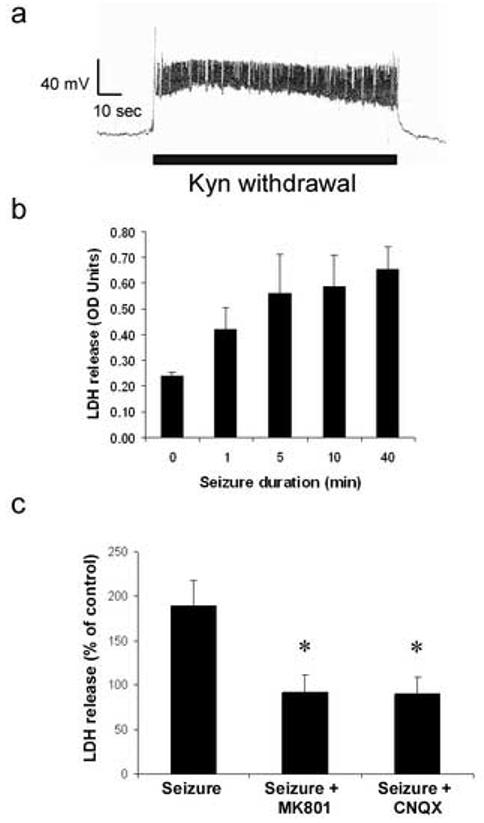
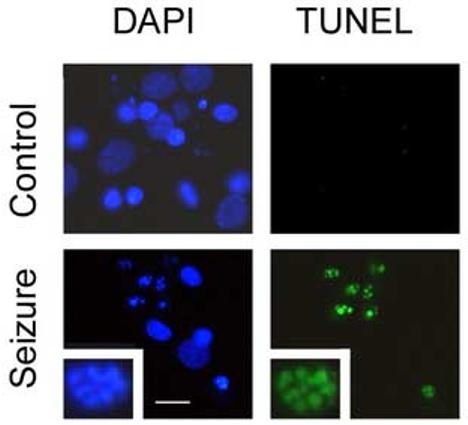
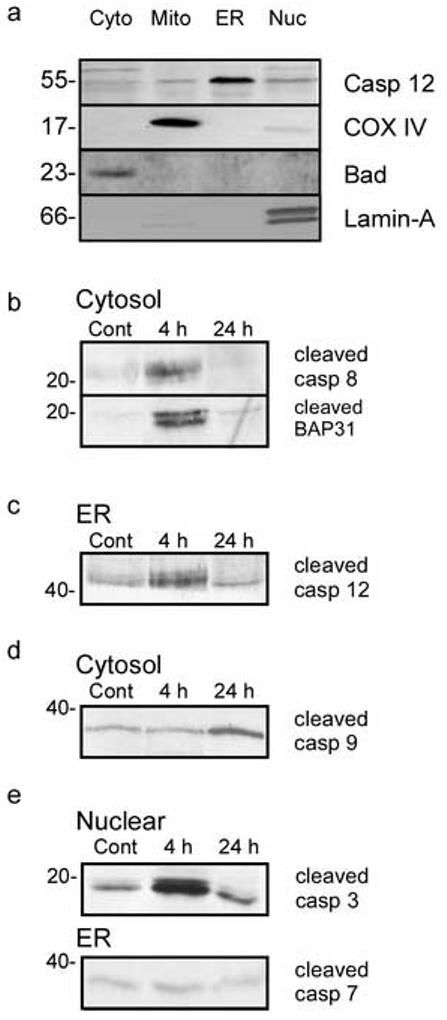
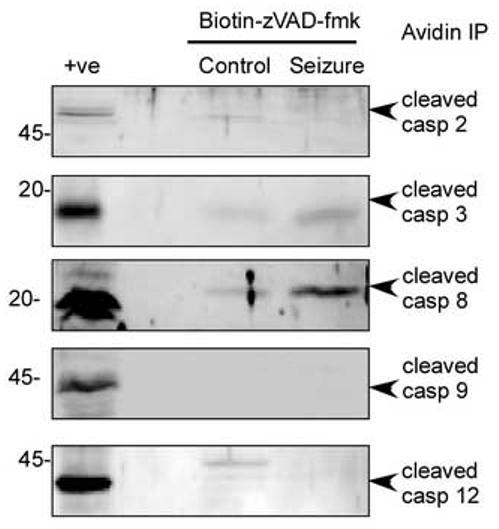
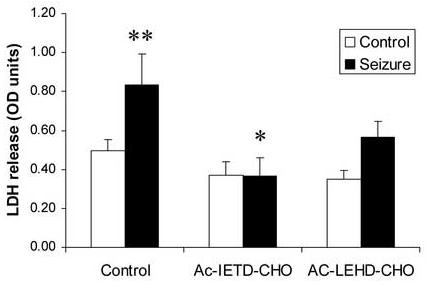
In response to harmful stresses, cells induce programmed cell death (PCD) or apoptosis. Seizures can induce neural damage and activate biochemical pathways associated with PCD. Since seizures trigger intracellular calcium overload, it has been presumed that the intrinsic cell death pathway mediated by mitochondrial dysfunction would modulate cell death following seizures. However, previous work suggests that the extrinsic cell death pathway may initiate the damage program. Here we investigate intrinsic versus extrinsic cell death pathway activation using caspase cleavage as a marker for activation of these pathways in a rat in vitro model of seizures. Hippocampal cells, chronically treated with kynurenic acid, had kynurenic acid withdrawn to induce seizure-like activity for 40 min. Subjecting rat hippocampal cultures to seizures increased cell death and apoptosis-like DNA fragmentation using TUNEL staining. Seizure-induced cell death was blocked by both MK801 (10 microM) and CNQX (40 microM), which suggests multiple glutamate receptors regulate seizure-induced cell death. Cleavage of the initiator caspases, caspase 8 and 12 were increased 4h following seizure, and cleavage of the quintessential executioner caspase, caspase 3 was increased 4h following seizure. In contrast, caspase 9 cleavage only increased 24h following seizure. Using an affinity labeling approach to trap activated caspases in situ, we show that caspase 8 is the apical caspase activated following seizures. Finally, we show that the caspase 8 inhibitor Ac-IETD-CHO was more effective at blocking seizure-induced cell death than the caspase 9 inhibitor Ac-LEHD-CHO. Taken together, our data suggests the extrinsic cell death pathway-associated caspase 8 is activated following seizures in vitro.
Figures
Similar articles
-
Seizure-like activity leads to the release of BAD from 14-3-3 protein and cell death in hippocampal neurons in vitro.Cell Death Differ. 2003 May;10(5):539-47. doi: 10.1038/sj.cdd.4401206. Cell Death Differ. 2003. PMID: 12728252
-
Cleavage of bid may amplify caspase-8-induced neuronal death following focally evoked limbic seizures.Neurobiol Dis. 2001 Aug;8(4):568-80. doi: 10.1006/nbdi.2001.0415. Neurobiol Dis. 2001. PMID: 11493022
-
Inhibition of caspase-8 attenuates neuronal death induced by limbic seizures in a cytochrome c-dependent and Smac/DIABLO-independent way.Brain Res. 2006 Jul 7;1098(1):204-11. doi: 10.1016/j.brainres.2006.04.131. Epub 2006 Jun 13. Brain Res. 2006. PMID: 16774749
-
Death and survival from executioner caspase activation.Semin Cell Dev Biol. 2024 Mar 15;156:66-73. doi: 10.1016/j.semcdb.2023.07.005. Epub 2023 Jul 18. Semin Cell Dev Biol. 2024. PMID: 37468421 Review.
-
Apoptosis and its Role in Parkinson’s Disease.In: Stoker TB, Greenland JC, editors. Parkinson’s Disease: Pathogenesis and Clinical Aspects [Internet]. Brisbane (AU): Codon Publications; 2018 Dec 21. Chapter 4. In: Stoker TB, Greenland JC, editors. Parkinson’s Disease: Pathogenesis and Clinical Aspects [Internet]. Brisbane (AU): Codon Publications; 2018 Dec 21. Chapter 4. PMID: 30702843 Free Books & Documents. Review.
Cited by
-
Neuroprotective Effects of Myricetin on PTZ-Induced Seizures in Mice: Evaluation of Oxidation, Neuroinflammation and Metabolism, and Apoptosis in the Hippocampus.Curr Issues Mol Biol. 2024 Aug 15;46(8):8914-8944. doi: 10.3390/cimb46080527. Curr Issues Mol Biol. 2024. PMID: 39194744 Free PMC article.
-
Caspases as master regulators of programmed cell death: apoptosis, pyroptosis and beyond.Exp Mol Med. 2025 Jun;57(6):1121-1132. doi: 10.1038/s12276-025-01470-9. Epub 2025 Jun 24. Exp Mol Med. 2025. PMID: 40555741 Free PMC article. Review.
-
KCC2 is required for the survival of mature neurons but not for their development.J Biol Chem. 2021 Jan-Jun;296:100364. doi: 10.1016/j.jbc.2021.100364. Epub 2021 Feb 2. J Biol Chem. 2021. PMID: 33539918 Free PMC article.
-
Mefloquine damage vestibular hair cells in organotypic cultures.Neurotox Res. 2011 Jul;20(1):51-8. doi: 10.1007/s12640-010-9221-z. Epub 2010 Sep 22. Neurotox Res. 2011. PMID: 20859773 Free PMC article.
-
Tolerance to ischemia - an increasingly complex biology.Transl Stroke Res. 2013 Feb;4(1):40-50. doi: 10.1007/s12975-012-0246-x. Epub 2013 Jan 11. Transl Stroke Res. 2013. PMID: 23504451 Free PMC article. Review.
References
-
- Ashkenazi A, Dixit VM. Death receptors: signaling and modulation. Science. 1998;281:1305–1308. - PubMed
-
- Blomgren K, Zhu C, Wang X, Karlsson JO, Leverin AL, Bahr BA, Mallard C, Hagberg H. Synergistic activation of caspase-3 by m-calpain after neonatal hypoxia-ischemia: a mechanism of “pathological apoptosis”? J. Biol. Chem. 2001;276:10191–10198. - PubMed
-
- Cao G, Minami M, Pei W, Yan C, Chen D, O'Horo C, Graham SH, Chen J. Intracellular bax translocation after transient cerebral ischemia: implications for a role of the mitochondrial apoptotic signaling pathway in ischemic neuronal death. J. Cereb. Blood Flow Metab. 2001;21:321–333. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
