

Dispersal is not female biased in a resource-defence mating ungulate, the European roe deer
- PMID: 16543177
- PMCID: PMC1560046
- DOI: 10.1098/rspb.2005.3329
Dispersal is not female biased in a resource-defence mating ungulate, the European roe deer
Abstract
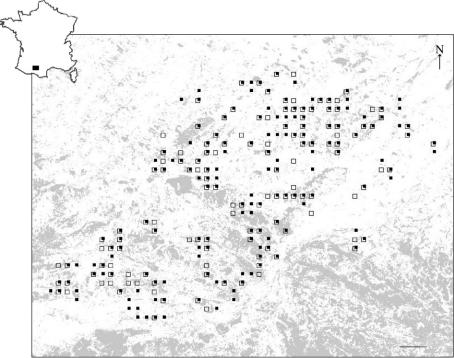
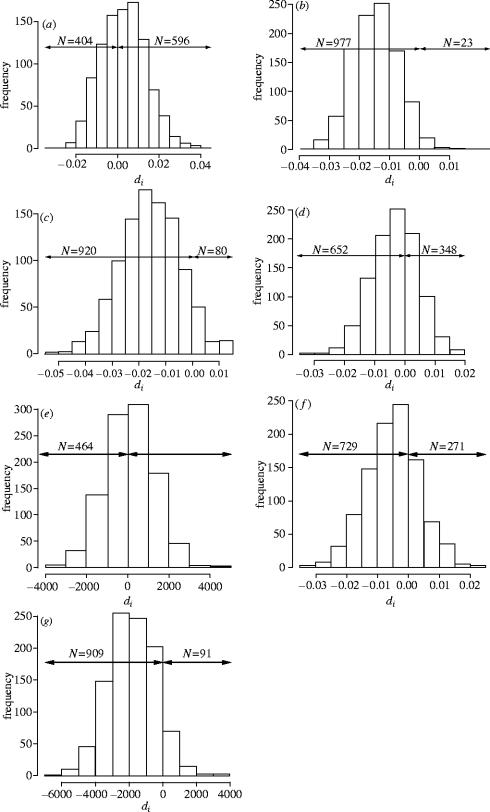
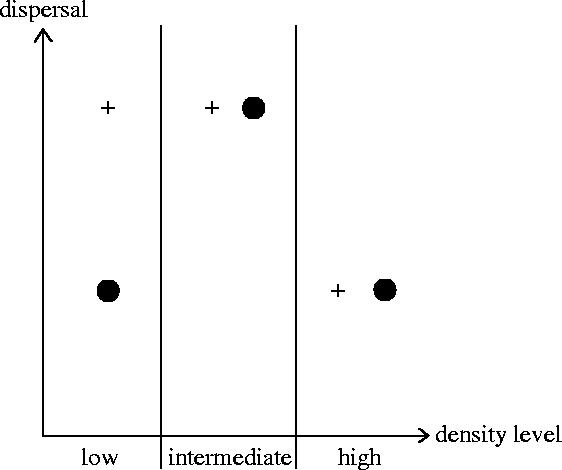
Dispersal is frequently more prevalent in one sex compared to the other. Greenwood proposed that patterns of sex-biased dispersal among birds and mammals are linked to their mating strategies. For species where males defend resources rather than females, he predicted female-biased dispersal, because males should remain at their birth site where they are familiar with the distribution of the resources that they must defend. Greenwood's hypothesis has been extensively supported among birds, where most species exhibit a resource-defence mating strategy. However, almost no equivalent information is available for mammals as males generally defend mates in this group. An exception is the European roe deer, a resource-defence mating ungulate. We thus tested Greenwood's hypothesis on this atypical mammalian model, looking for female-biased dispersal using sex-specific inter-individual genetic distances. We conclusively show that gene flow is not higher among females compared to males in the studied roe deer population, and hence that dispersal is not female-biased, suggesting that male mating strategy is not the primary selective force driving the evolution of dispersal in roe deer. We discuss the role of female mate choice and intra-sexual competition as possible alternative selective pressures involved.
Figures
References
-
- Austin J.D, Davila J.A, Lougheed S.C, Boag P.T. Genetic evidence for female-biased dispersal in the bullfrog, Rana catesbeiana (Ranidae) Mol. Ecol. 2003;12:3165–3172. doi:10.1046/j.1365-294X.2003.01948.x - DOI - PubMed
-
- Cargnelutti B, Reby D, Desneux L, Angibault J.M, Joachim J, Hewison A.J.M. Space use by roe deer in a fragmented landscape. Some preliminary results. Rev. Ecol. (Terre Vie) 2002;57:29–37.
-
- Coulon A, Cosson J.F, Angibault J.M, Cargnelutti B, Galan M, Morellet N, Petit E, Aulagnier S, Hewison A.J.M. Landscape connectivity influences gene flow in a roe deer population inhabiting a fragmented landscape: an individual-based approach. Mol. Ecol. 2004;13:2841–2850. doi:10.1111/j.1365-294X.2004.02253.x - DOI - PubMed
-
- Cransac N, Cibien C, Angibault J.M, Morellet N, Vincent J.P, Hewison A.J.M. Variations saisonnières du régime alimentaire du chevreuil (Capreolus capreolus) selon le sexe en milieu forestier à forte densité (forêt domaniale de Dourdan) Mammalia. 2001;65:1–12.
-
- Danilkin A, Hewison A.J.M. Wildlife Ecology and Behavioural Series. Chapman & Hall; London: 1996. Behavioural ecology of Siberian and European Roe Deer.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
