

Telomeric heterochromatin boundaries require NuA4-dependent acetylation of histone variant H2A.Z in Saccharomyces cerevisiae
- PMID: 16543222
- PMCID: PMC1413290
- DOI: 10.1101/gad.1386306
Telomeric heterochromatin boundaries require NuA4-dependent acetylation of histone variant H2A.Z in Saccharomyces cerevisiae
Abstract
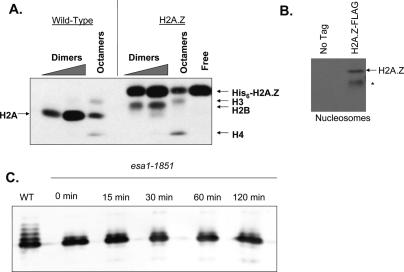
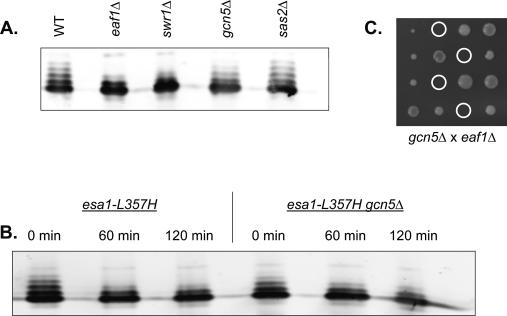
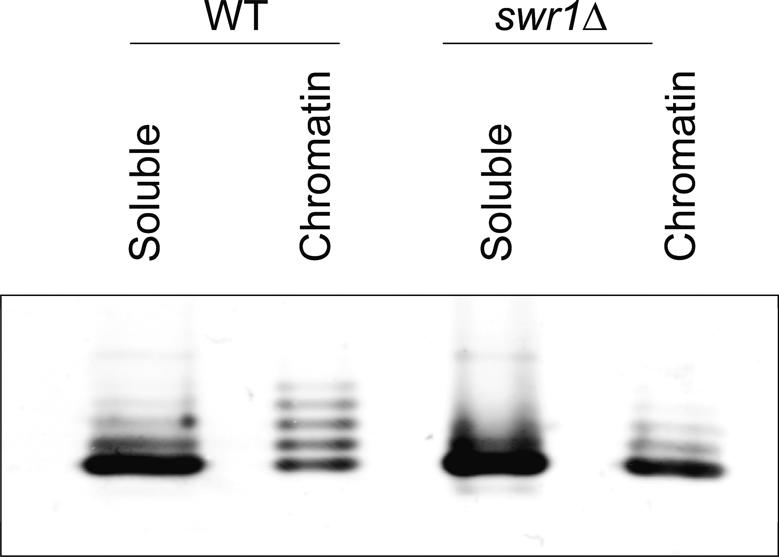
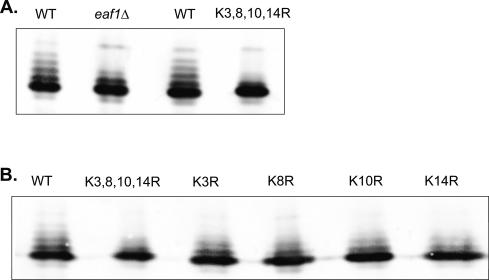
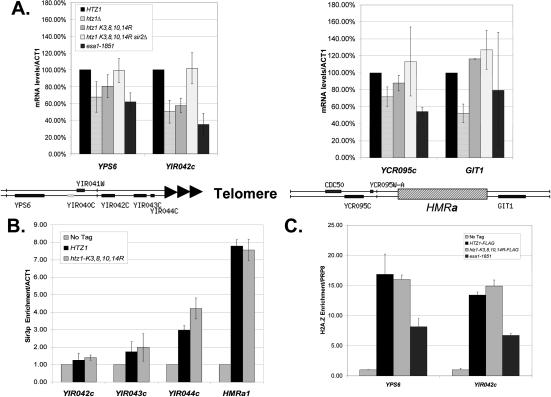
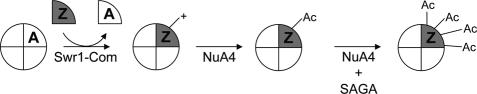
SWR1-Com, which is responsible for depositing H2A.Z into chromatin, shares four subunits with the NuA4 histone acetyltransferase complex. This overlap in composition led us to test whether H2A.Z was a substrate of NuA4 in vitro and in vivo. The N-terminal tail of H2A.Z was acetylated in vivo at multiple sites by a combination of NuA4 and SAGA. H2A.Z acetylation was also dependent on SWR1-Com, causing H2A.Z to be efficiently acetylated only when incorporated in chromatin. Unacetylatable H2A.Z mutants were, like wild-type H2A.Z, enriched at heterochromatin boundaries, but were unable to block spreading of heterochromatin. A mutant version of H2A.Z that could not be acetylated, in combination with a mutation in a nonessential gene in the NuA4 complex, caused a pronounced decrease in growth rate. This H2A.Z mutation was lethal in combination with a mutant version of histone H4 that could not be acetylated by NuA4. Taken together, these results show a role for H2A.Z acetylation in restricting silent chromatin, and reveal that acetylation of H2A.Z and H4 can contribute to a common function essential to life.
Figures

References
-
- Bird A.W., Yu D.Y., Pray-Grant M.G., Qiu Q., Harmon K.E., Megee P.C., Grant P.A., Smith M.M., Christman M.F. Acetylation of histone H4 by Esa1 is required for DNA double-strand break repair. Nature. 2002;419:411–415. - PubMed
-
- Carmen A.A., Milne L., Grunstein M. Acetylation of the yeast histone H4 N terminus regulates its binding to heterochromatin protein SIR3. J. Biol. Chem. 2002;277:4778–4781. - PubMed
-
- Chakravarthy S., Bao Y., Roberts V.A., Tremethick D., Luger K. Structural characterization of histone H2A variants. Cold Spring Harb. Symp. Quant. Biol. 2004;69:227–234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases