

Carbonic anhydrase and its influence on carbon isotope discrimination during C4 photosynthesis. Insights from antisense RNA in Flaveria bidentis
- PMID: 16543411
- PMCID: PMC1459309
- DOI: 10.1104/pp.106.077776
Carbonic anhydrase and its influence on carbon isotope discrimination during C4 photosynthesis. Insights from antisense RNA in Flaveria bidentis
Abstract
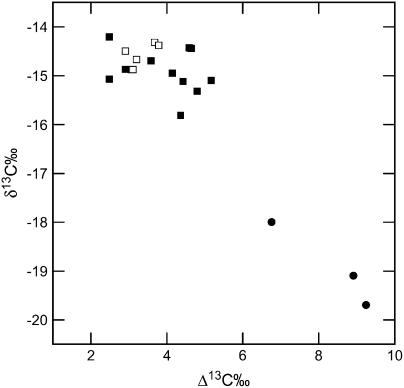
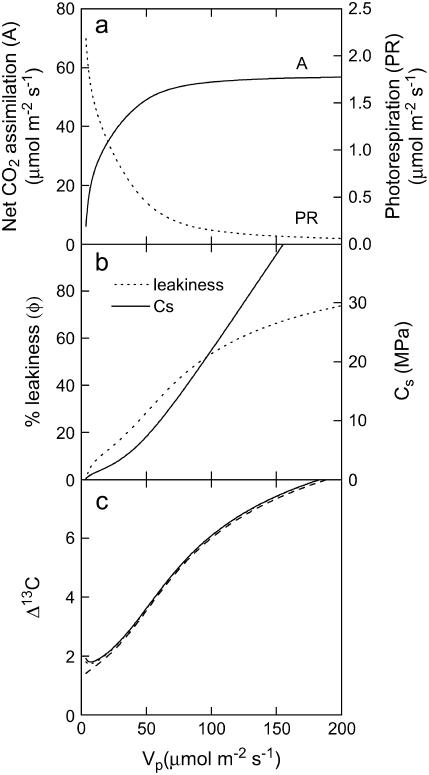
In C4 plants, carbonic anhydrase (CA) facilitates both the chemical and isotopic equilibration of atmospheric CO2 and bicarbonate (HCO3-) in the mesophyll cytoplasm. The CA-catalyzed reaction is essential for C4 photosynthesis, and the model of carbon isotope discrimination (Delta13C) in C4 plants predicts that changes in CA activity will influence Delta13C. However, experimentally, the influence of CA on Delta13C has not been demonstrated in C4 plants. Here, we compared measurements of Delta13C during C4 photosynthesis in Flaveria bidentis wild-type plants with F. bidentis plants with reduced levels of CA due to the expression of antisense constructs targeted to a putative mesophyll cytosolic CA. Plants with reduced CA activity had greater Delta13C, which was also evident in the leaf dry matter carbon isotope composition (delta13C). Contrary to the isotope measurements, photosynthetic rates were not affected until CA activity was less than 20% of wild type. Measurements of Delta13C, delta13C of leaf dry matter, and rates of net CO2 assimilation were all dramatically altered when CA activity was less than 5% of wild type. CA activity in wild-type F. bidentis is sufficient to maintain net CO2 assimilation; however, reducing leaf CA activity has a relatively large influence on Delta13C, often without changes in net CO2 assimilation. Our data indicate that the extent of CA activity in C4 leaves needs to be taken into account when using Delta13C and/or delta13C to model the response of C4 photosynthesis to changing environmental conditions.
Figures
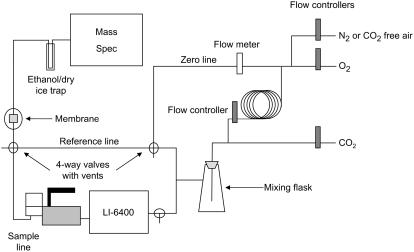
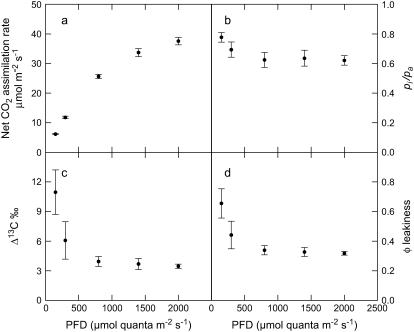
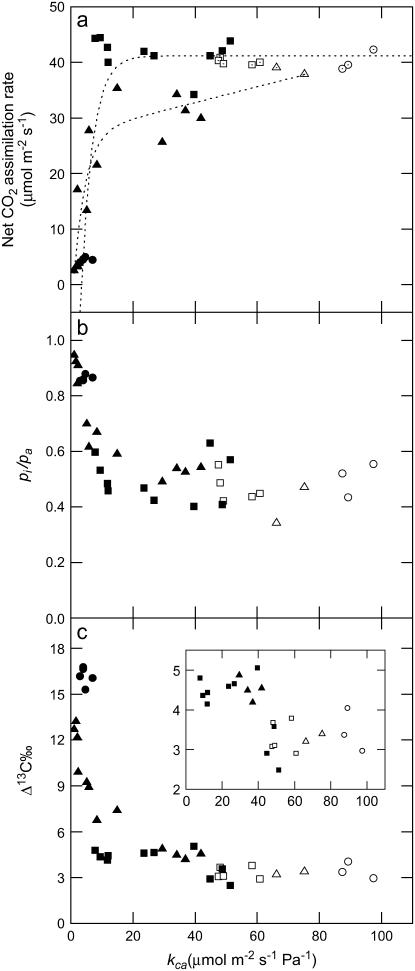
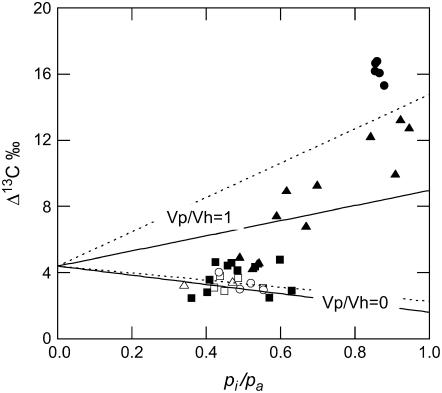
References
-
- Ainsworth EA, Long SP (2005) What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy. New Phytol 165: 351–371 - PubMed
-
- Berry JA, Farquhar GD (1978) The CO2-concentrating function of C4 photosynthesis. A biochemical model. In DO Hall, J Coombs, TW Goodwin, eds, Proceedings of the Fourth International Congress on Photosynthesis. Biochemical Society, London, pp 119–131
-
- Brownell P, Bielig L, Grof C (1991) Increased carbonic anhydrase activity in leaves of sodium-deficient C4 plants. Aust J Plant Physiol 18: 589–592
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
