

Pertactin beta-helix folding mechanism suggests common themes for the secretion and folding of autotransporter proteins
- PMID: 16549796
- PMCID: PMC1458770
- DOI: 10.1073/pnas.0507923103
Pertactin beta-helix folding mechanism suggests common themes for the secretion and folding of autotransporter proteins
Abstract
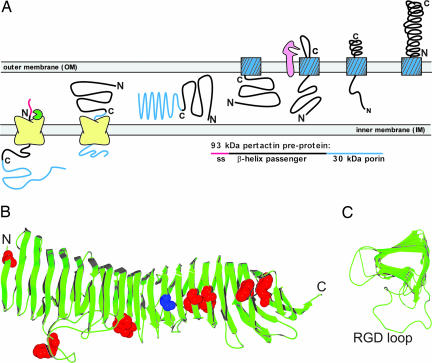
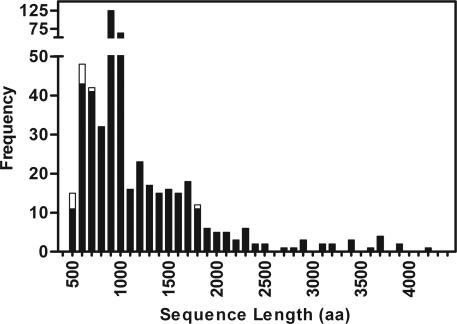
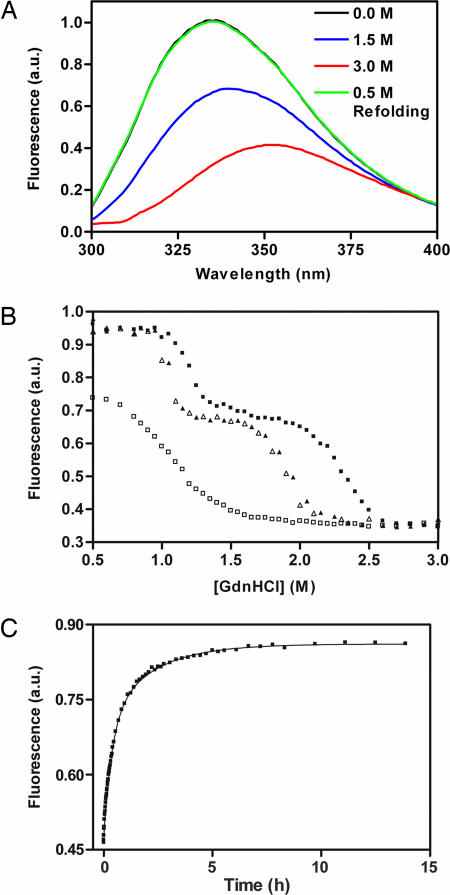
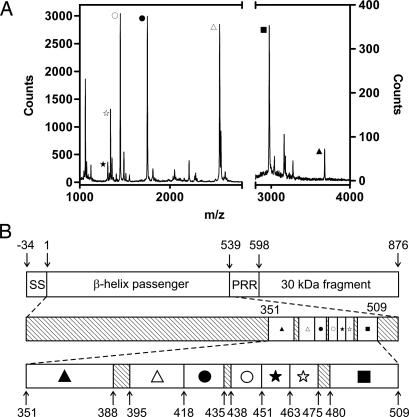
Many virulence factors secreted from pathogenic Gram-negative bacteria are autotransporter proteins. The final step of autotransporter secretion is C --> N-terminal threading of the passenger domain through the outer membrane (OM), mediated by a cotranslated C-terminal porin domain. The native structure is formed only after this final secretion step, which requires neither ATP nor a proton gradient. Sequence analysis reveals that, despite size, sequence, and functional diversity among autotransporter passenger domains, >97% are predicted to form parallel beta-helices, indicating this structural topology may be important for secretion. We report the folding behavior of pertactin, an autotransporter passenger domain from Bordetella pertussis. The pertactin beta-helix folds reversibly in isolation, but folding is much slower than expected based on size and native-state topology. Surprisingly, pertactin is not prone to aggregation during folding, even though folding is extremely slow. Interestingly, equilibrium denaturation results in the formation of a partially folded structure, a stable core comprising the C-terminal half of the protein. Examination of the pertactin crystal structure does not reveal any obvious reason for the enhanced stability of the C terminus. In vivo, slow folding would prevent premature folding of the passenger domain in the periplasm, before OM secretion. Moreover, the extra stability of the C-terminal rungs of the beta-helix might serve as a template for the formation of native protein during OM secretion; hence, vectorial folding of the beta-helix could contribute to the energy-independent translocation mechanism. Coupled with the sequence analysis, the results presented here suggest a general mechanism for autotransporter secretion.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures

References
-
- Skillman K. M., Barnard T. J., Peterson J. H., Ghirlando R., Bernstein H. D. Mol. Microbiol. 2005;58:945–958. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
