

A murine Mll-AF4 knock-in model results in lymphoid and myeloid deregulation and hematologic malignancy
- PMID: 16551973
- PMCID: PMC1895483
- DOI: 10.1182/blood-2005-08-3498
A murine Mll-AF4 knock-in model results in lymphoid and myeloid deregulation and hematologic malignancy
Abstract
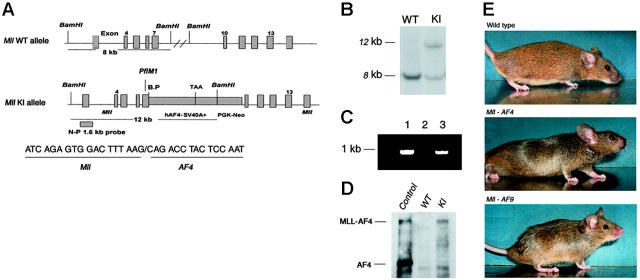
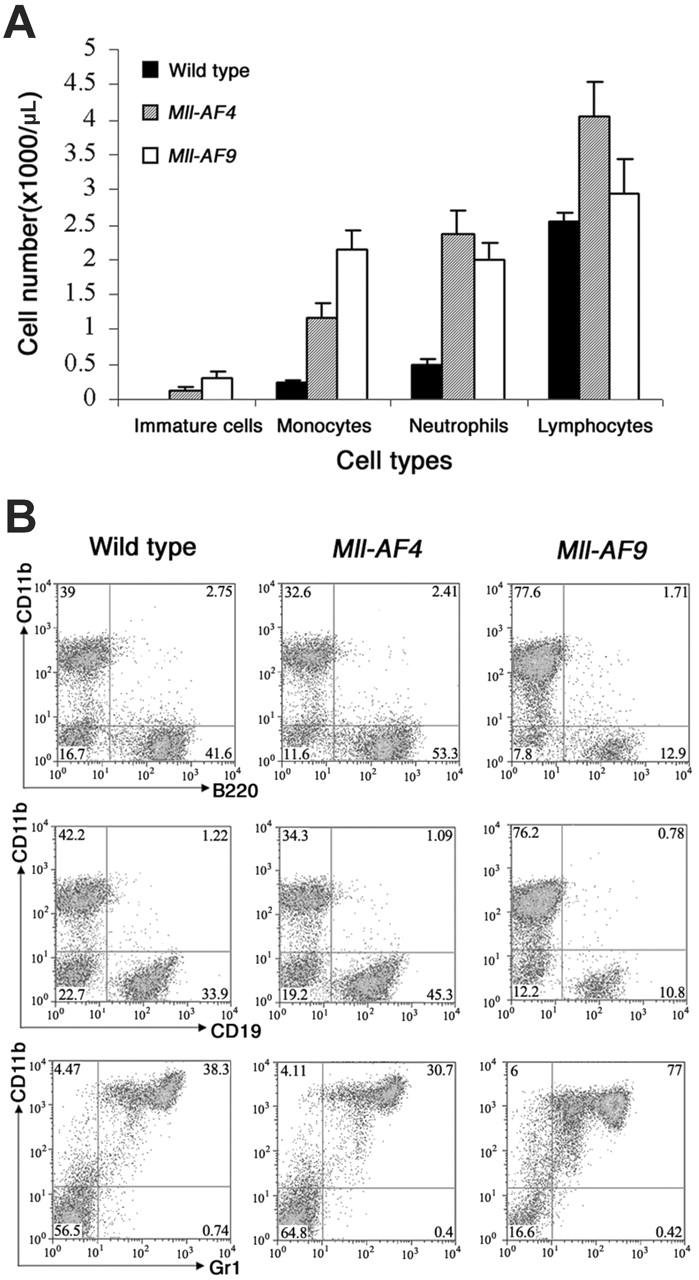
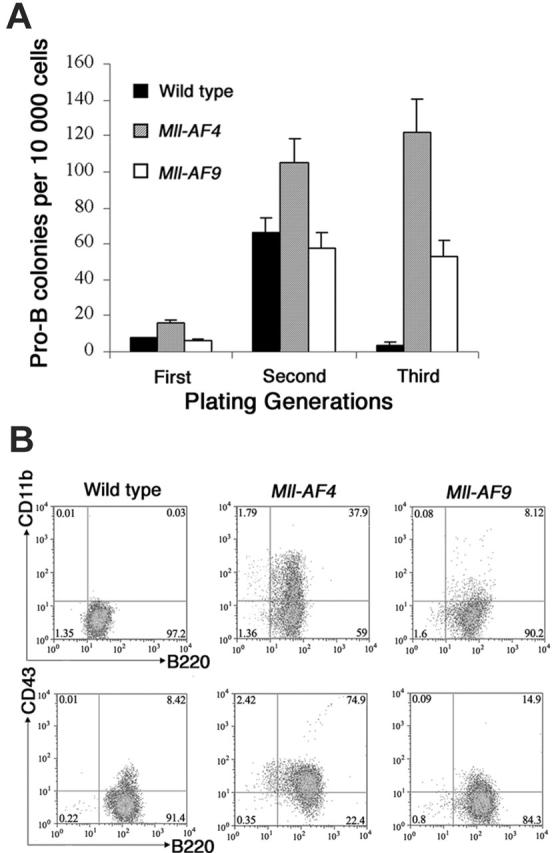
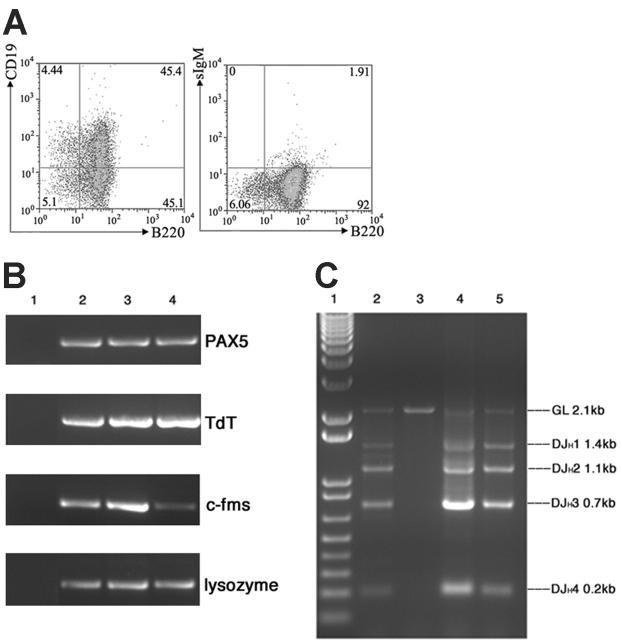
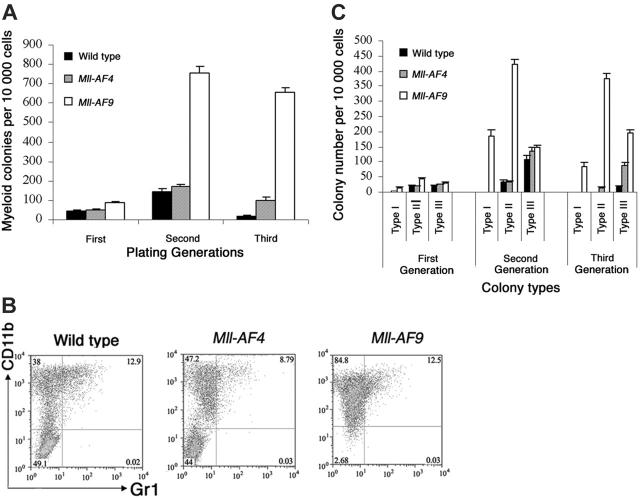
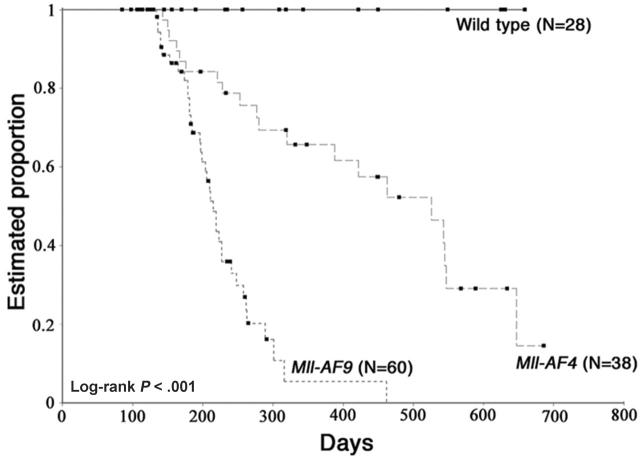
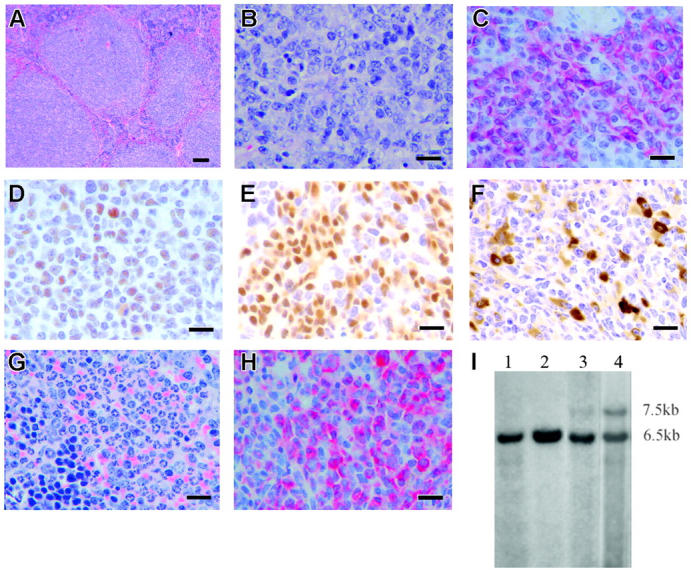
The 2 most frequent human MLL hematopoietic malignancies involve either AF4 or AF9 as fusion partners; each has distinct biology but the role of the fusion partner is not clear. We produced Mll-AF4 knock-in (KI) mice by homologous recombination in embryonic stem cells and compared them with Mll-AF9 KI mice. Young Mll-AF4 mice had lymphoid and myeloid deregulation manifest by increased lymphoid and myeloid cells in hematopoietic organs. In vitro, bone marrow cells from young mice formed unique mixed pro-B lymphoid (B220(+)CD19(+)CD43(+)sIgM(-), PAX5(+), TdT(+), IgH rearranged)/myeloid (CD11b/Mac1(+), c-fms(+), lysozyme(+)) colonies when grown in IL-7- and Flt3 ligand-containing media. Mixed lymphoid/myeloid hyperplasia and hematologic malignancies (most frequently B-cell lymphomas) developed in Mll-AF4 mice after prolonged latency; long latency to malignancy indicates that Mll-AF4-induced lymphoid/myeloid deregulation alone is insufficient to produce malignancy. In contrast, young Mll-AF9 mice had predominately myeloid deregulation in vivo and in vitro and developed myeloid malignancies. The early onset of distinct mixed lymphoid/myeloid lineage deregulation in Mll-AF4 mice shows evidence for both "instructive" and "noninstructive" roles for AF4 and AF9 as partners in MLL fusion genes. The molecular basis for "instruction" and secondary cooperating mutations can now be studied in our Mll-AF4 model.
Figures
References
-
- Ayton PM, Cleary ML. Molecular mechanisms of leukemogenesis mediated by MLL fusion proteins. Oncogene. 2001;20: 5695-5707. - PubMed
-
- Armstrong SA, Staunton JE, Silverman LB, et al. MLL translocations specify a distinct gene expression profile that distinguishes a unique leukemia. Nat Genet. 2002;30: 41-47. - PubMed
-
- Chen CS, Sorensen PH, Domer PH, et al. Molecular rearrangements on chromosome 11q23 predominate in infant acute lymphoblastic leukemia and are associated with specific biologic variables and poor outcome. Blood. 1993;81: 2386-2393. - PubMed
-
- Dimartino JF, Cleary ML. Mll rearrangements in haematological malignancies: lessons from clinical and biological studies. Br J Haematol. 1999;106: 614-626. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
