

Analysis of involvement of the RecF pathway in p44 recombination in Anaplasma phagocytophilum and in Escherichia coli by using a plasmid carrying the p44 expression and p44 donor loci
- PMID: 16552034
- PMCID: PMC1418890
- DOI: 10.1128/IAI.74.4.2052-2062.2006
Analysis of involvement of the RecF pathway in p44 recombination in Anaplasma phagocytophilum and in Escherichia coli by using a plasmid carrying the p44 expression and p44 donor loci
Abstract
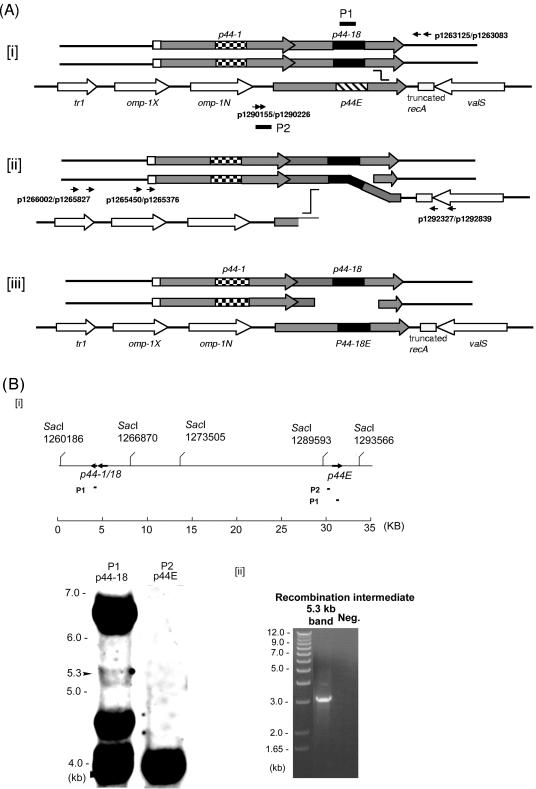
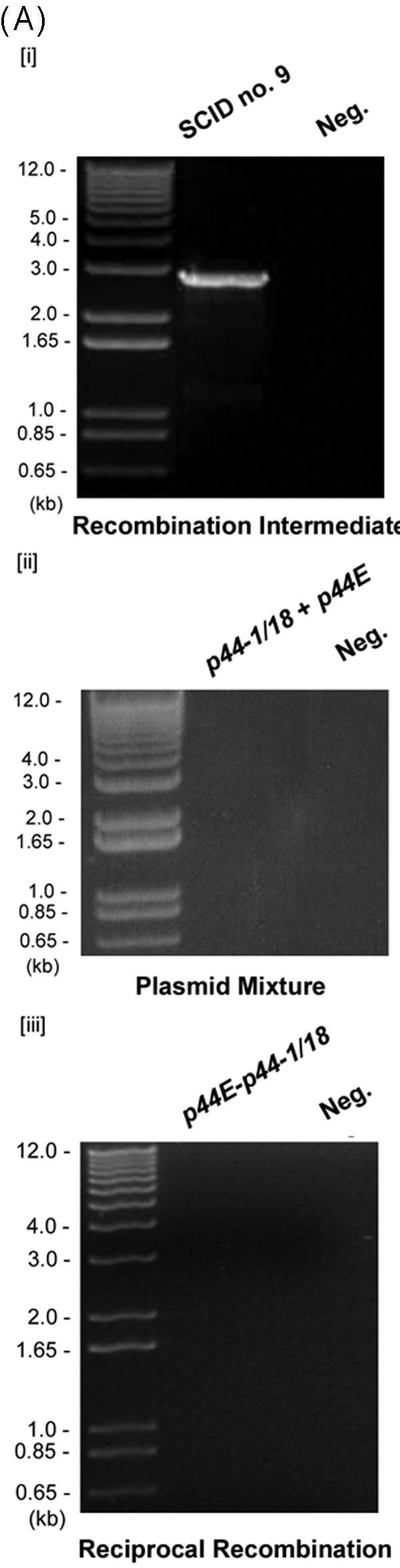
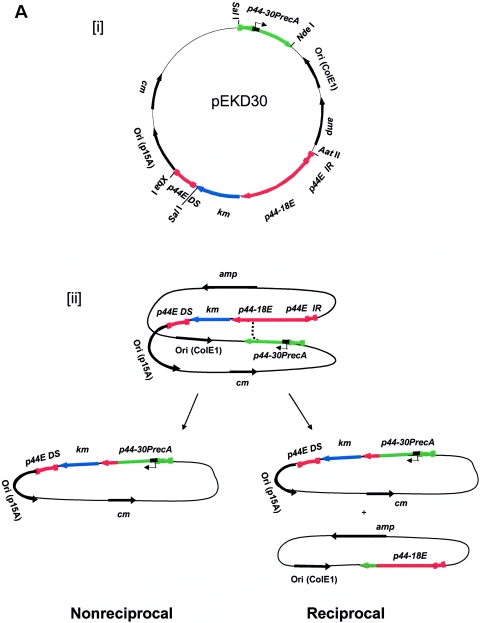
Anaplasma phagocytophilum, the etiologic agent of human granulocytic anaplasmosis, has a large paralog cluster (approximate 90 members) that encodes the 44-kDa major outer membrane proteins (P44s). Gene conversion at a single p44 expression locus leads to P44 antigenic variation. Homologs of genes for the RecA-dependent RecF pathway, but not the RecBCD or RecE pathways, of recombination were detected in the A. phagocytophilum genome. In the present study, we examined whether the RecF pathway is involved in p44 gene conversion. The recombination intermediate structure between a donor p44 and the p44 expression locus of A. phagocytophilum was detected in an HL-60 cell culture by Southern blot analysis followed by sequencing the band and in blood samples from infected SCID mice by PCR, followed by sequencing. The sequences were consistent with the RecF pathway recombination: a half-crossover structure, consisting of the donor p44 locus connected to the 3' conserved region of the recipient p44 in the p44 expression locus in direct orientation. To determine whether the p44 recombination intermediate structure can be generated in a RecF-active Escherichia coli strain, we constructed a double-origin plasmid carrying the p44 expression locus and a donor p44 locus and introduced the plasmid into various E. coli strains. The recombination intermediate was recovered in an E. coli strain with active RecF recombination pathway but not in strains with deficient RecF pathway. Our results support the view that the p44 gene conversion in A. phagocytophilum occurs through the RecF pathway.
Figures


References
-
- Allers, T., and M. Lichten. 2001. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106:47-57. - PubMed
-
- Bakken, J. S., J. Krueth, C. Wilson-Nordskog, R. L. Tilden, K. Asanovich, and J. S. Dumler. 1996. Clinical and laboratory characteristics of human granulocytic ehrlichiosis. JAMA 275:199-205. - PubMed
-
- Barbet, A. F., P. F. M. Meeus, M. Belanger, M. V. Bowie, J. Yi, A. M. Lundgren, A. R. Alleman, S. J. Wong, F. K. Chu, U. G. Munderloh, and S. D. Jauron. 2003. Expression of multiple outer membrane protein sequence variants from a single genomic locus of Anaplasma phagocytophilum. Infect. Immun. 71:1706-1718. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
