

NF-kappa B-inducing kinase regulates selected gene expression in the Nod2 signaling pathway
- PMID: 16552041
- PMCID: PMC1418900
- DOI: 10.1128/IAI.74.4.2121-2127.2006
NF-kappa B-inducing kinase regulates selected gene expression in the Nod2 signaling pathway
Abstract
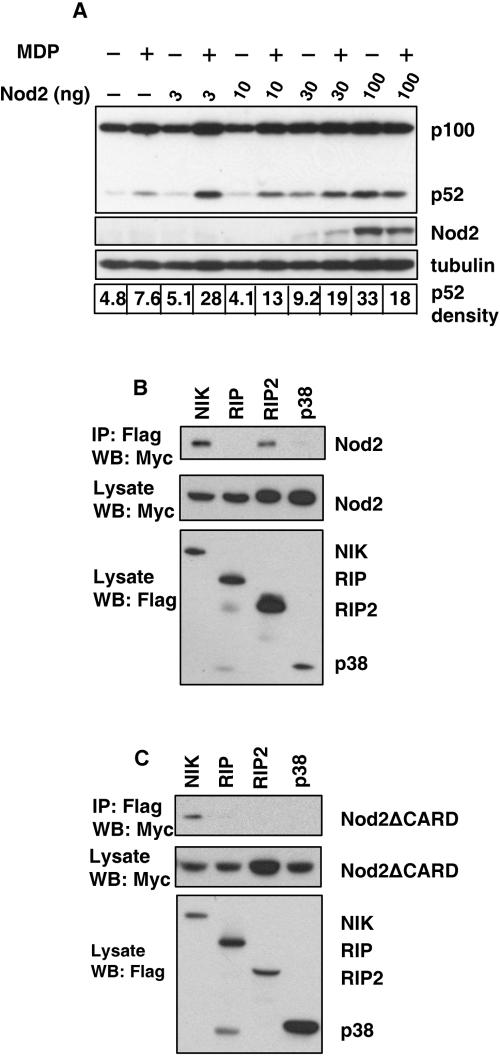
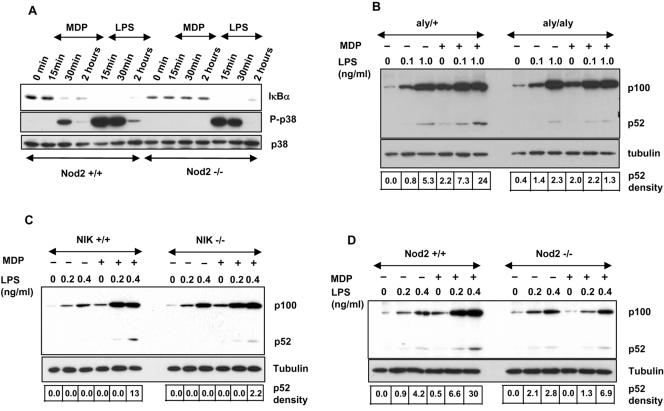
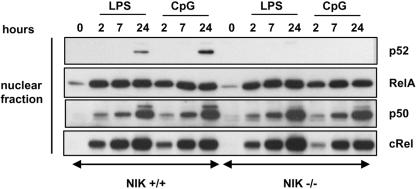
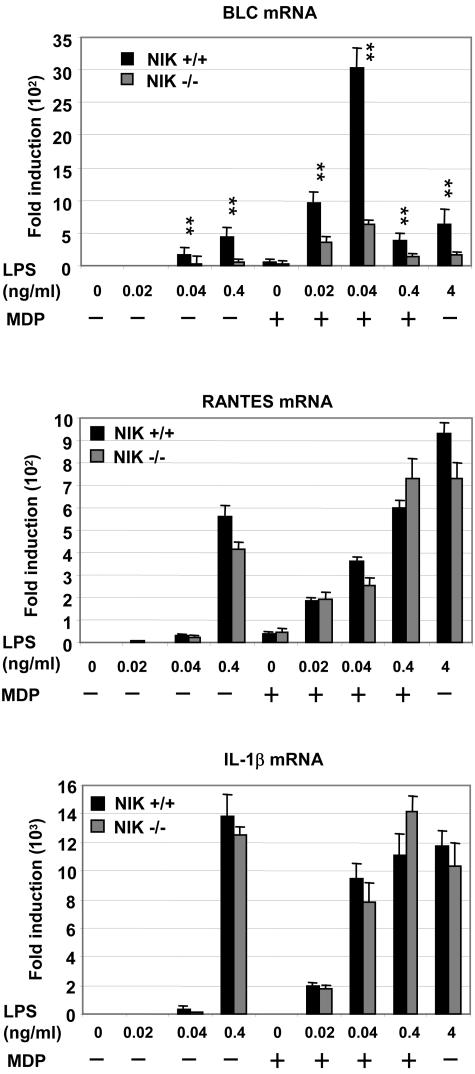
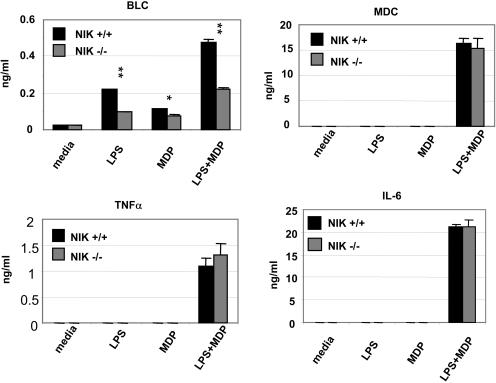
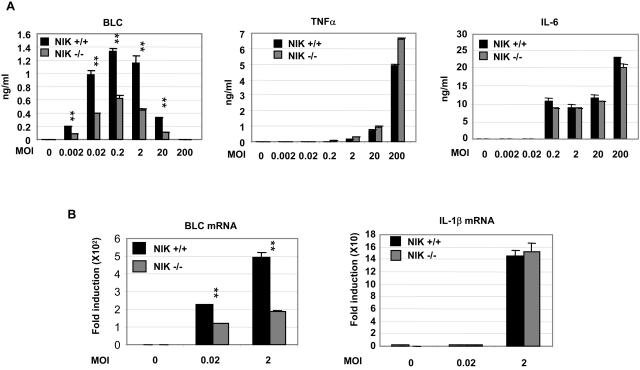
The innate immune system surveys the extra- and intracellular environment for the presence of microbes. Among the intracellular sensors is a protein known as Nod2, a cytosolic protein containing a leucine-rich repeat domain. Nod2 is believed to play a role in determining host responses to invasive bacteria. A key element in upregulating host defense involves activation of the NF-kappaB pathway. It has been suggested through indirect studies that NF-kappaB-inducing kinase, or NIK, may be involved in Nod2 signaling. Here we have used macrophages derived from primary explants of bone marrow from wild-type mice and mice that either bear a mutation in NIK, rendering it inactive, or are derived from NIK-/- mice, in which the NIK gene has been deleted. We show that NIK binds to Nod2 and mediates induction of specific changes induced by the specific Nod2 activator, muramyl dipeptide, and that the role of NIK occurs in settings where both the Nod2 and TLR4 pathways are activated by their respective agonists. Specifically, we have linked NIK to the induction of the B-cell chemoattractant known as BLC and suggest that this chemokine may play a role in processes initiated by Nod2 activation that lead to improved host defense.
Figures
References
-
- Akira, S., and K. Takeda. 2004. Toll-like receptor signalling. Nat. Rev. Immunol. 4:499-511. - PubMed
-
- Beutler, B., K. Hoebe, X. Du, and R. J. Ulevitch. 2003. How we detect microbes and respond to them: the Toll-like receptors and their transducers. J. Leukoc. Biol. 74:479-485. - PubMed
-
- Beutler, B., and J. Hoffmann. 2004. Innate immunity. Curr. Opin. Immunol. 16:1-3. - PubMed
-
- Bonizzi, G., and M. Karin. 2004. The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 25:280-288. - PubMed
-
- Carlsen, H. S., E. S. Baekkevold, H. C. Morton, G. Haraldsen, and P. Brandtzaeg. 2004. Monocyte-like and mature macrophages produce CXCL13 (B cell-attracting chemokine 1) in inflammatory lesions with lymphoid neogenesis. Blood 104:3021-3027. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
