

Tau protects microtubules in the axon from severing by katanin
- PMID: 16554463
- PMCID: PMC6674103
- DOI: 10.1523/JNEUROSCI.5392-05.2006
Tau protects microtubules in the axon from severing by katanin
Abstract
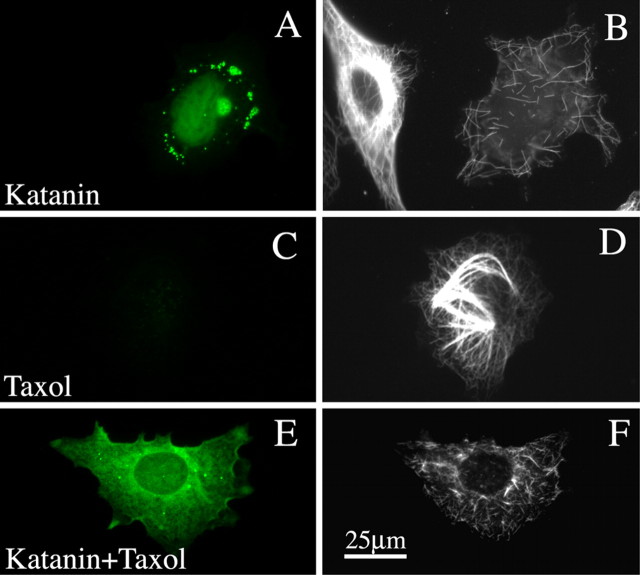
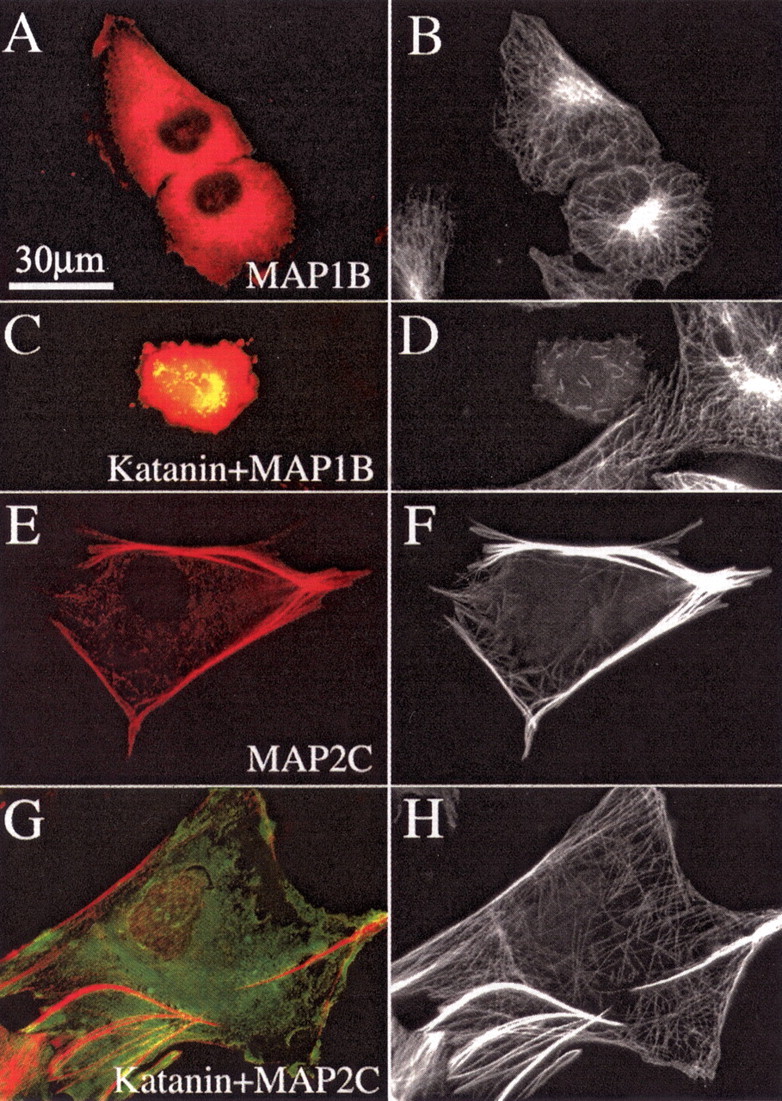
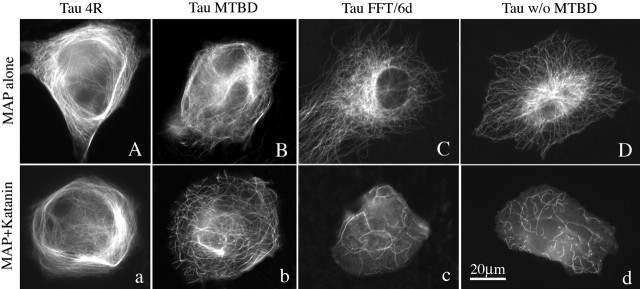
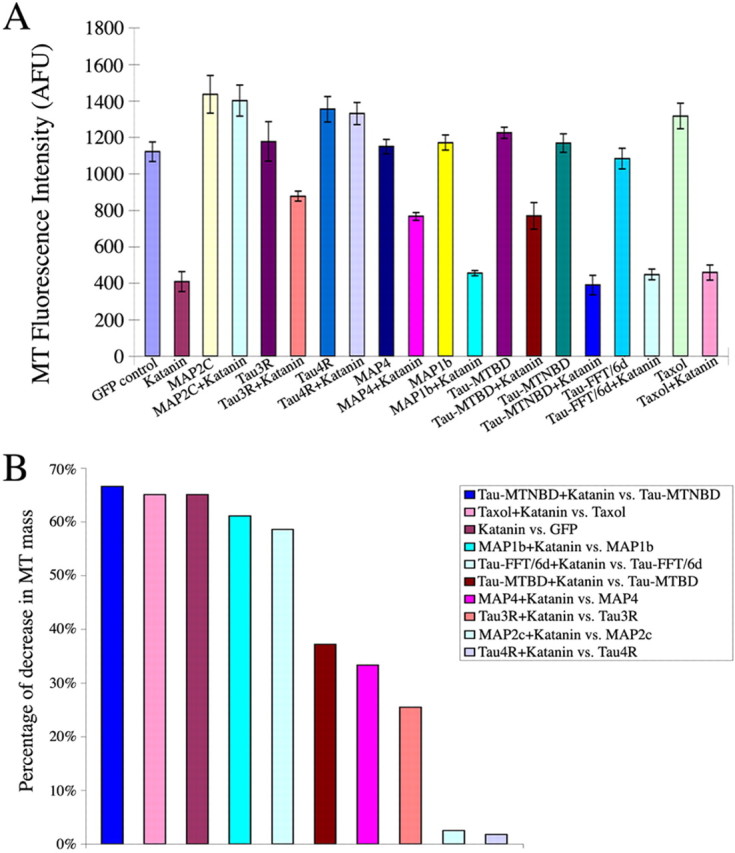
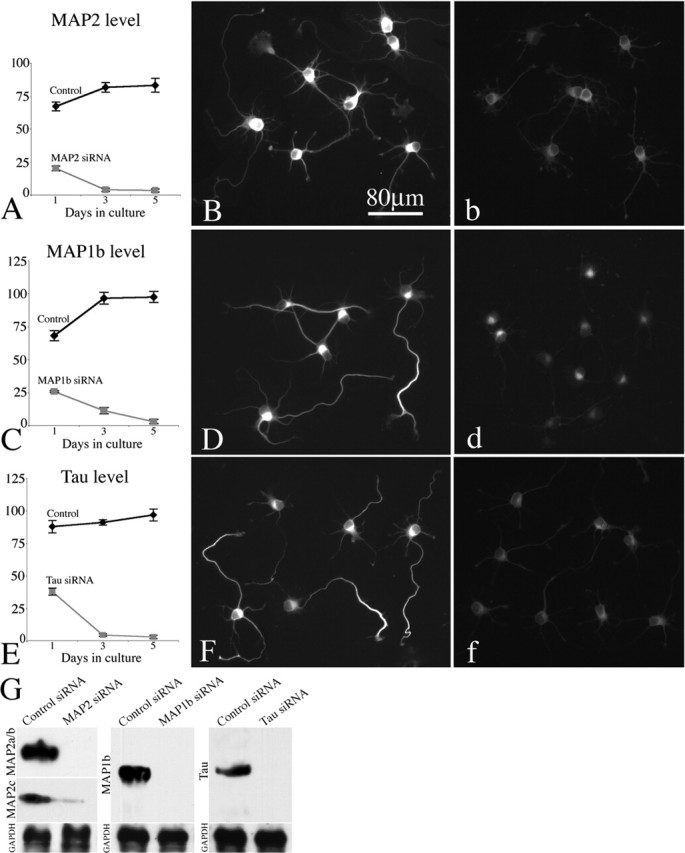
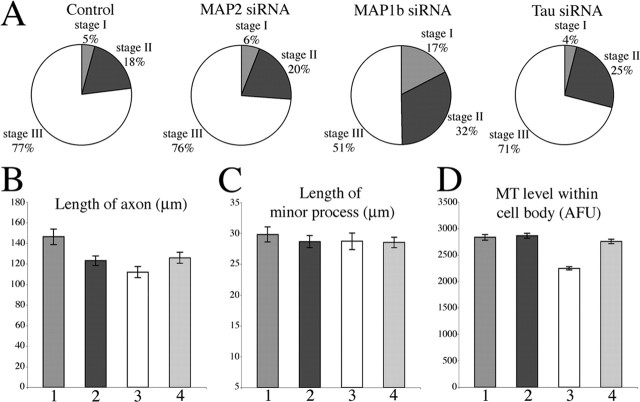
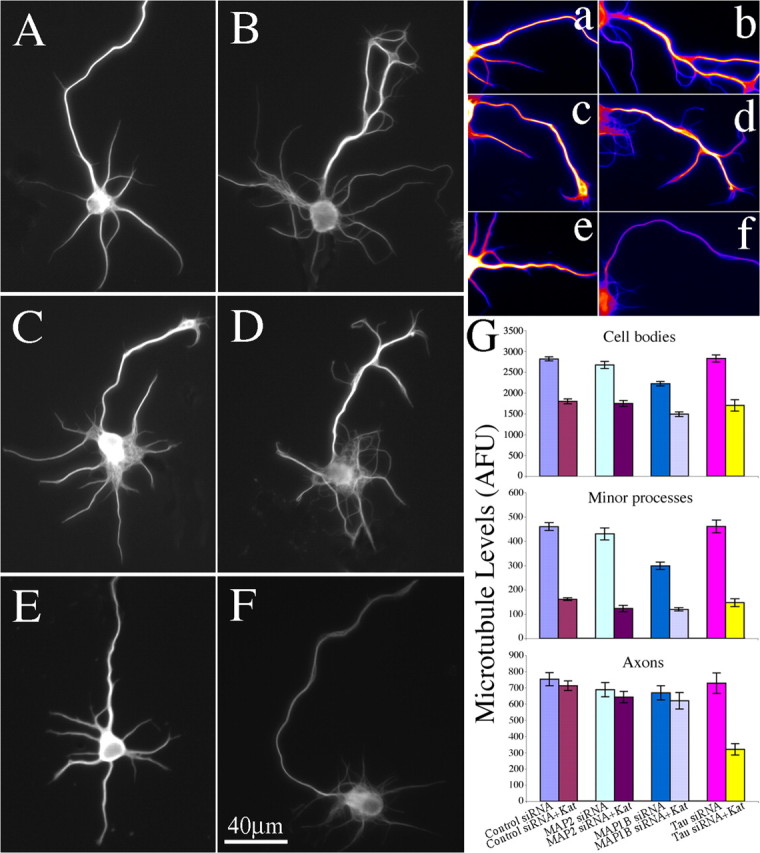
Microtubules in the axon are more resistant to severing by katanin than microtubules elsewhere in the neuron. We have hypothesized that this is because of the presence of tau on axonal microtubules. When katanin is overexpressed in fibroblasts, the microtubules are severed into short pieces, but this phenomenon is suppressed by the coexpression of tau. Protection against severing is also afforded by microtubule-associated protein 2 (MAP2), which has a tau-like microtubule-binding domain, but not by MAP1b, which has a different microtubule-binding domain. The microtubule-binding domain of tau is required for the protection, but within itself, provides less protection than the entire molecule. When tau (but not MAP2 or MAP1b) is experimentally depleted from neurons, the microtubules in the axon lose their characteristic resistance to katanin. These results, which validate our hypothesis, also suggest a potential explanation for why axonal microtubules deteriorate in neuropathies involving the dissociation of tau from the microtubules.
Figures
References
-
- Baas PW, Ahmad FJ (2001). Force generation by cytoskeletal motor proteins as a regulator of axonal elongation and retraction. Trends Cell Biol 11:244–249. - PubMed
-
- Baas PW, Qiang L (2005). Neuronal microtubules: when the MAP is the roadblock. Trends Cell Biol 15:183–187. - PubMed
-
- Brugg B, Reddy D, Matus A (1993). Attenuation of microtubule-associated protein 1B expression by antisense oligodeoxynucleotides inhibits initiation of neurite outgrowth. Neuroscience 52:489–496. - PubMed
-
- Caceres A, Kosik KS (1990). Inhibition of neurite polarity by tau antisense oligonucleotides in primary cerebellar neurons. Nature 343:461–463. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials