

Motoneurons derived from embryonic stem cells express transcription factors and develop phenotypes characteristic of medial motor column neurons
- PMID: 16554476
- PMCID: PMC6674087
- DOI: 10.1523/JNEUROSCI.5537-05.2006
Motoneurons derived from embryonic stem cells express transcription factors and develop phenotypes characteristic of medial motor column neurons
Abstract
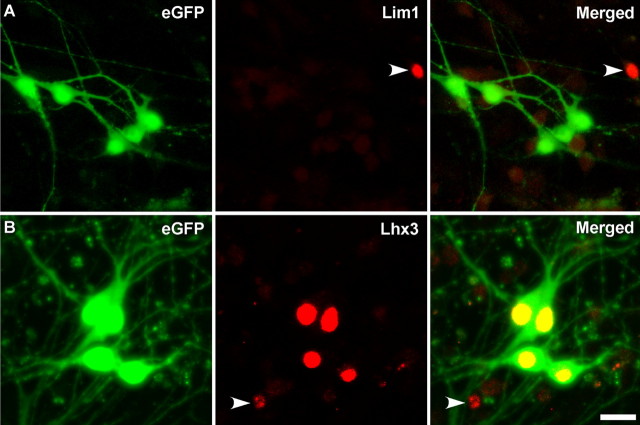
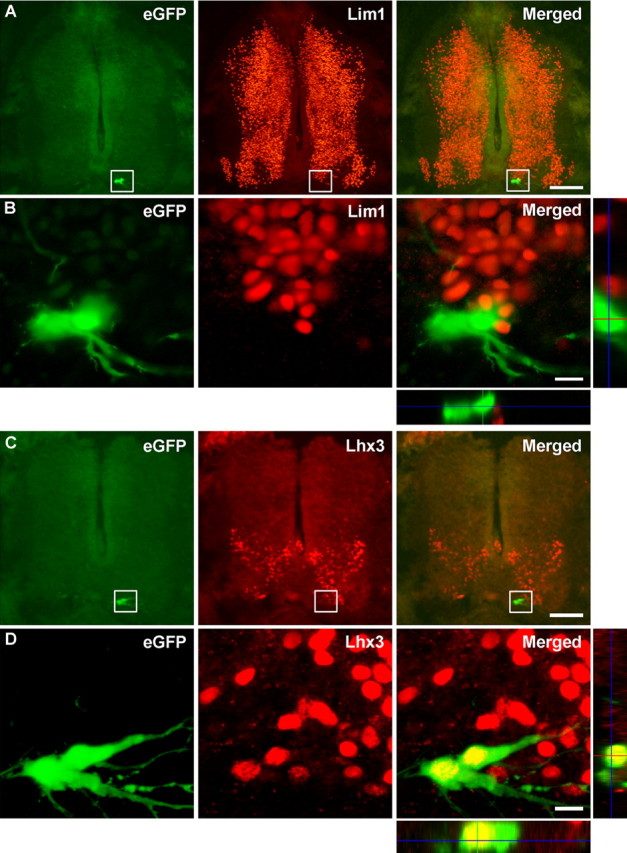
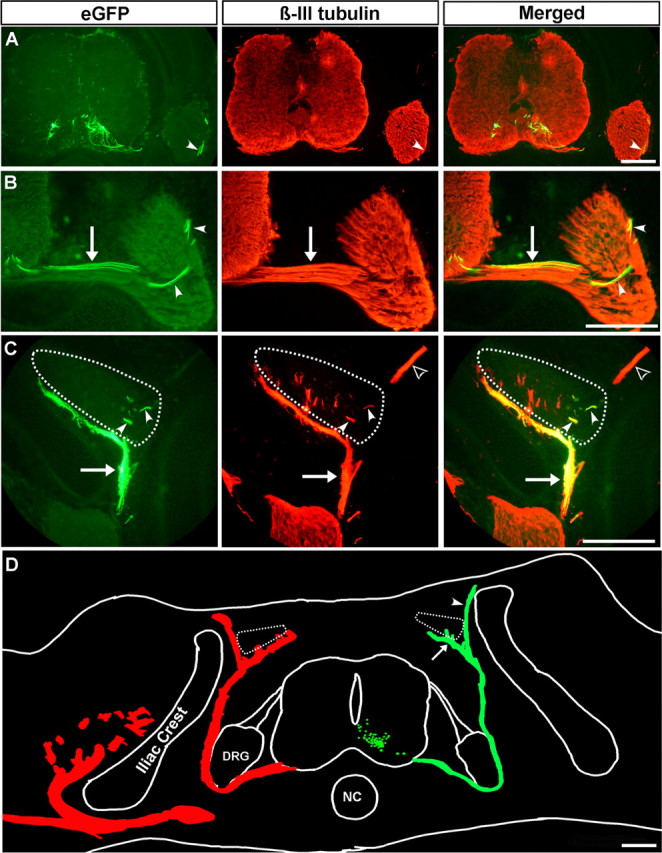
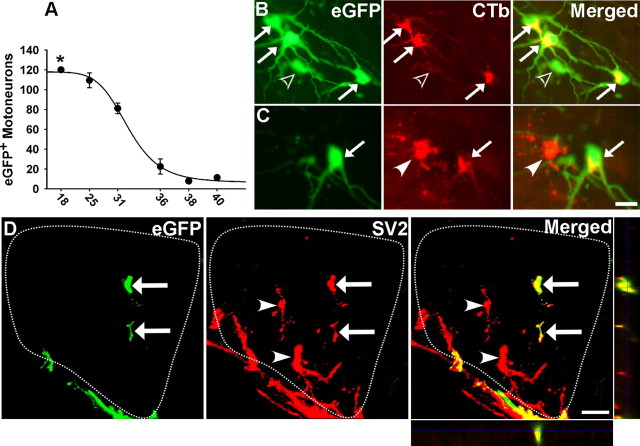
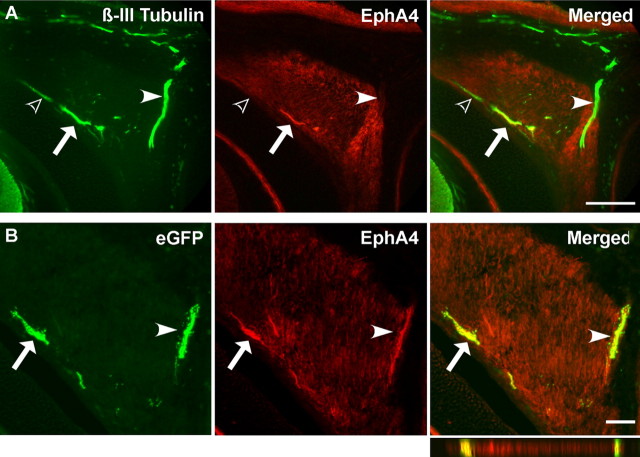
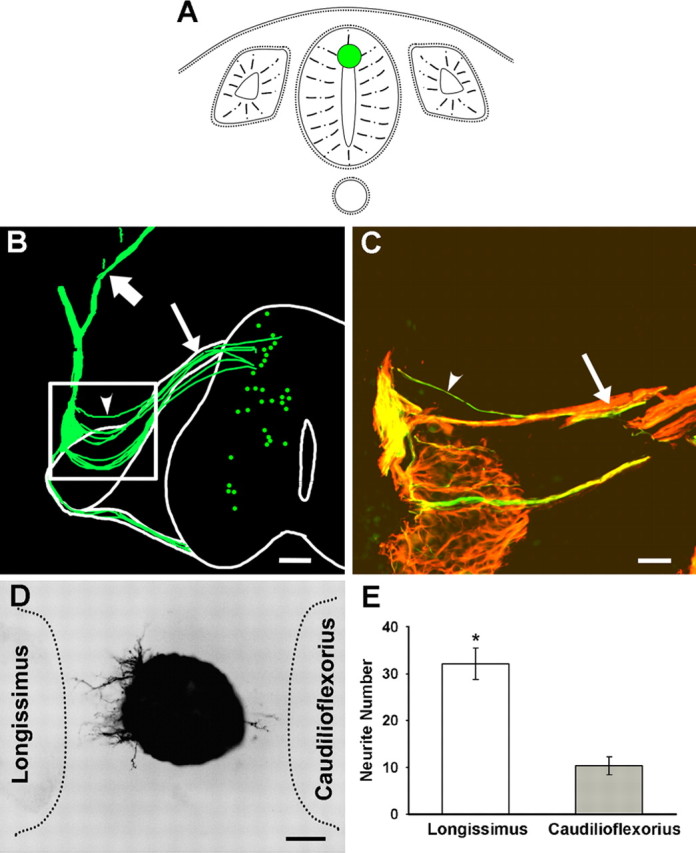
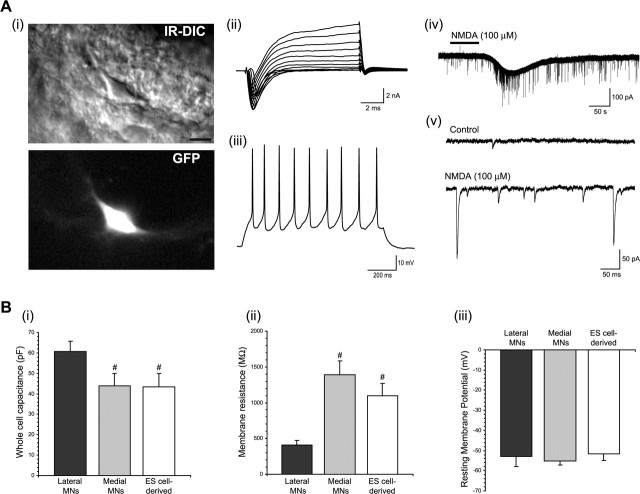
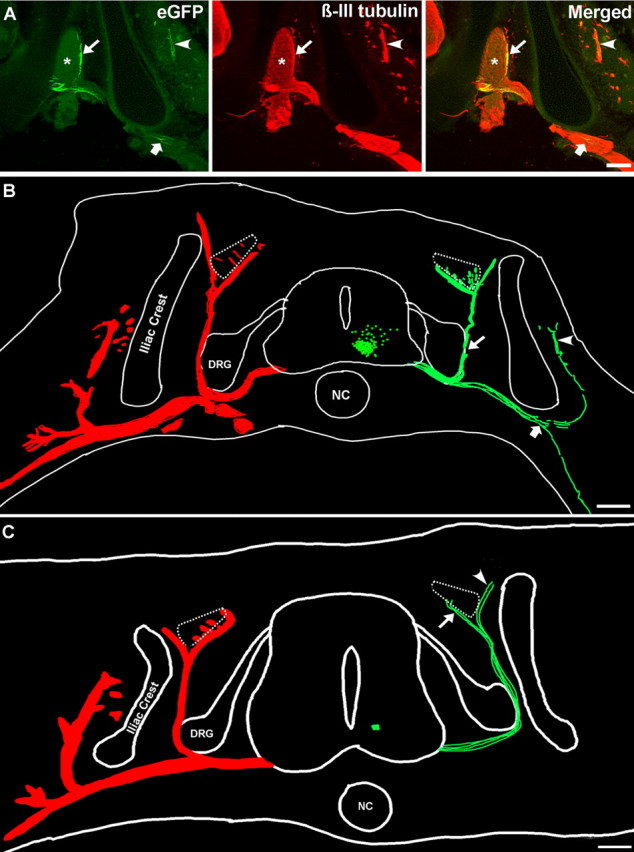
Embryonic stem (ES) cells differentiate into functional motoneurons when treated with a sonic hedgehog (Shh) agonist and retinoic acid (RA). Whether ES cells can be directed to differentiate into specific subtypes of motoneurons is unknown. We treated embryoid bodies generated from HBG3 ES cells with a Shh agonist and RA for 5 d in culture to induce motoneuron differentiation. Enhanced green fluorescent protein (eGFP) expression was used to identify putative motoneurons, because eGFP is expressed under the control of the Hb9 promoter in HBG3 cells. We found that 96 +/- 0.7% of the differentiated eGFP+ motoneurons expressed Lhx3, a homeobox gene expressed by postmitotic motoneurons in the medial motor column (MMCm), when the treated cells were plated on a neurite-promoting substrate for 5 d. When the treated embryoid bodies were transplanted into stage 17 chick neural tubes, the eGFP+ motoneurons migrated to the MMCm, expressed Lhx3, projected axons to the appropriate target for MMCm motoneurons (i.e., epaxial muscles), and contained synaptic vesicles within intramuscular axonal branches. In ovo and in vitro studies indicated that chemotropic factors emanating from the epaxial muscle and/or surrounding mesenchyme likely guide Lhx3+ motoneurons to their correct target. Finally, whole-cell patch-clamp recordings of transplanted ES cell-derived motoneurons demonstrated that they received synaptic input, elicited repetitive trains of action potentials, and developed passive membrane properties that were similar to host MMCm motoneurons. These results indicate that ES cells can be directed to form subtypes of neurons with specific phenotypic properties.
Figures
References
-
- Abercrombie M (1946). Estimation of nuclear population from microtome sections. Anat Neurobiol Rec 94:239–247. - PubMed
-
- Bjorklund LM, Isacson O (2002). Regulation of dopamine cell type and transmitter function in fetal and stem cell transplantation for Parkinson's disease. Prog Brain Res 138:411–420. - PubMed
-
- Chisholm A, Tessier-Lavigne M (1999). Conservation and divergence of axon guidance mechanisms. Curr Opin Neurobiol 9:603–615. - PubMed
-
- Dasen JS, Tice BC, Brenner-Morton S, Jessell TM (2005). A hox regulatory network establishes motor neuron pool identity and target-muscle connectivity. Cell 123:477–491. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials