

CHIP-mediated stress recovery by sequential ubiquitination of substrates and Hsp70
- PMID: 16554822
- PMCID: PMC4112096
- DOI: 10.1038/nature04600
CHIP-mediated stress recovery by sequential ubiquitination of substrates and Hsp70
Abstract
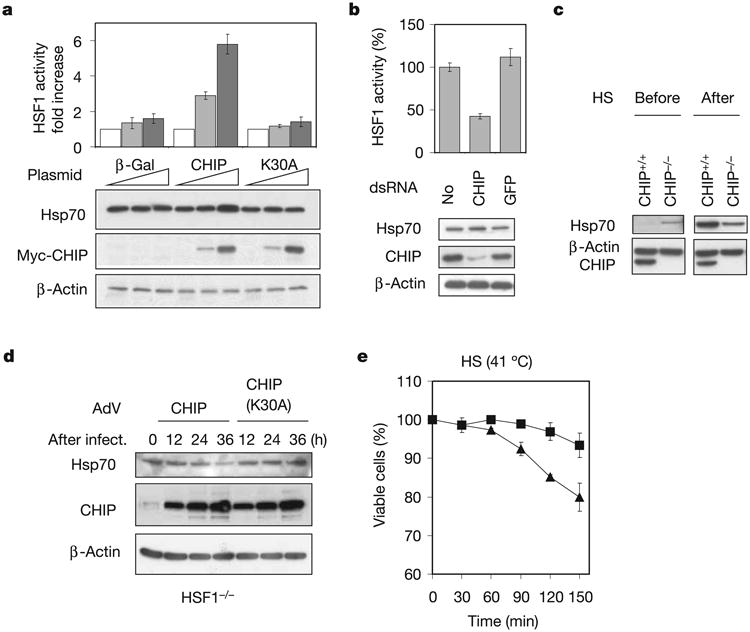
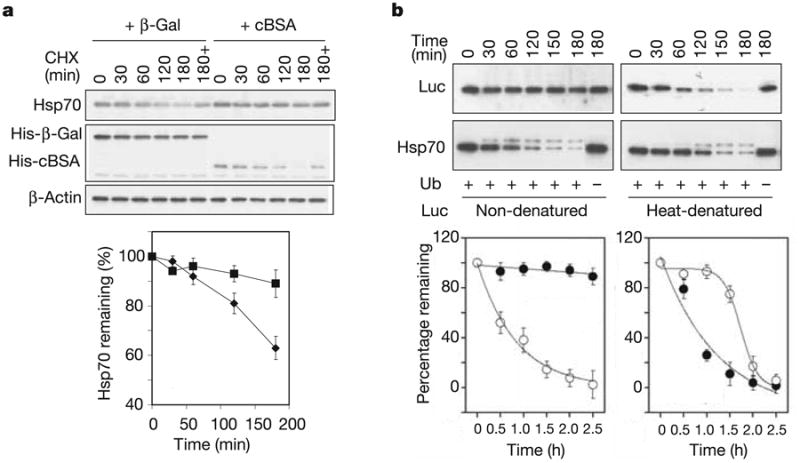
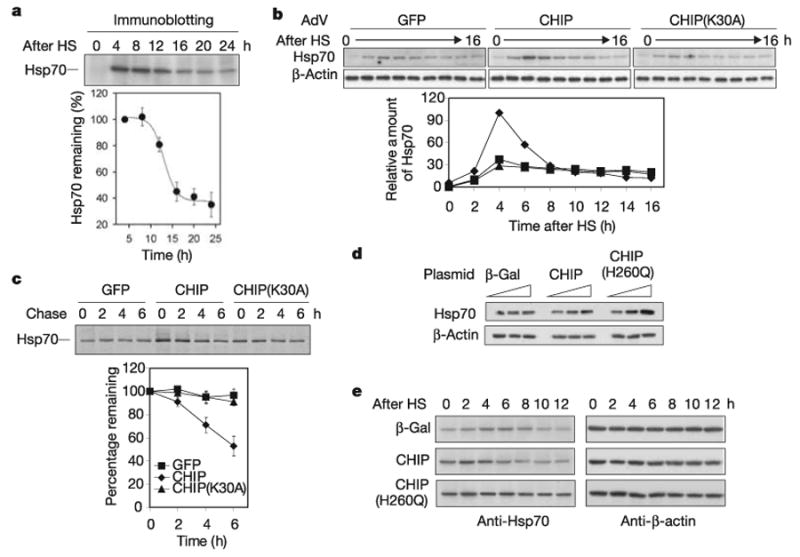
Exposure of cells to various stresses often leads to the induction of a group of proteins called heat shock proteins (HSPs, molecular chaperones). Hsp70 is one of the most highly inducible molecular chaperones, but its expression must be maintained at low levels under physiological conditions to permit constitutive cellular activities to proceed. Heat shock transcription factor 1 (HSF1) is the transcriptional regulator of HSP gene expression, but it remains poorly understood how newly synthesized HSPs return to basal levels when HSF1 activity is attenuated. CHIP (carboxy terminus of Hsp70-binding protein), a dual-function co-chaperone/ubiquitin ligase, targets a broad range of chaperone substrates for proteasomal degradation. Here we show that CHIP not only enhances Hsp70 induction during acute stress but also mediates its turnover during the stress recovery process. Central to this dual-phase regulation is its substrate dependence: CHIP preferentially ubiquitinates chaperone-bound substrates, whereas degradation of Hsp70 by CHIP-dependent targeting to the ubiquitin-proteasome system occurs when misfolded substrates have been depleted. The sequential catalysis of the CHIP-associated chaperone adaptor and its bound substrate provides an elegant mechanism for maintaining homeostasis by tuning chaperone levels appropriately to reflect the status of protein folding within the cytoplasm.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
-
- Parsell DA, Lindquist S. The function of heat-shock proteins in stress tolerance: degradation and reactivation of damaged proteins. Annu Rev Genet. 1993;27:437–496. - PubMed
-
- Morimoto RI. Cells in stress: transcriptional activation of heat shock genes. Science. 1993;259:1409–1410. - PubMed
-
- Feder JH, Rossi JM, Solomon J, Solomon N, Lindquist S. The consequences of expressing hsp70 in Drosophila cells at normal temperatures. Genes Dev. 1992;6:1402–1413. - PubMed
-
- Volloch VZ, Sherman MY. Oncogenic potential of Hsp72. Oncogene. 1999;18:3648–3651. - PubMed
-
- Wu C. Heat shock transcription factors: structure and regulation. Annu Rev Cell Dev Biol. 1995;11:441–469. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
