

Isl1Cre reveals a common Bmp pathway in heart and limb development
- PMID: 16556916
- PMCID: PMC5576437
- DOI: 10.1242/dev.02322
Isl1Cre reveals a common Bmp pathway in heart and limb development
Abstract
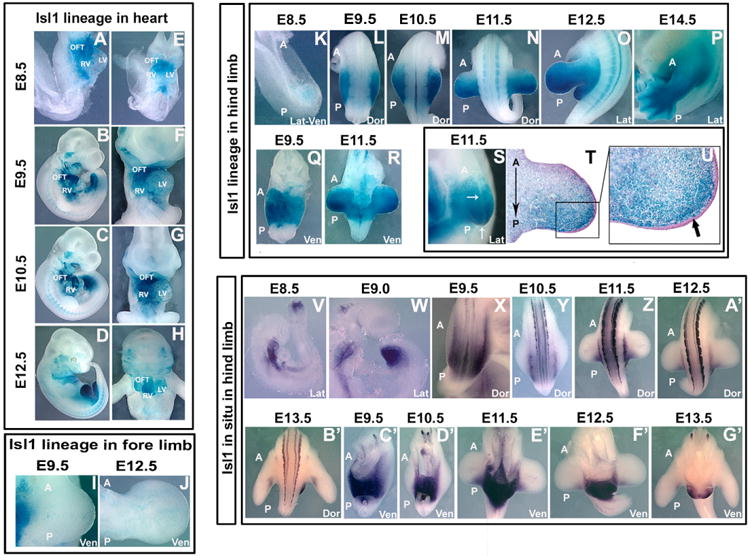
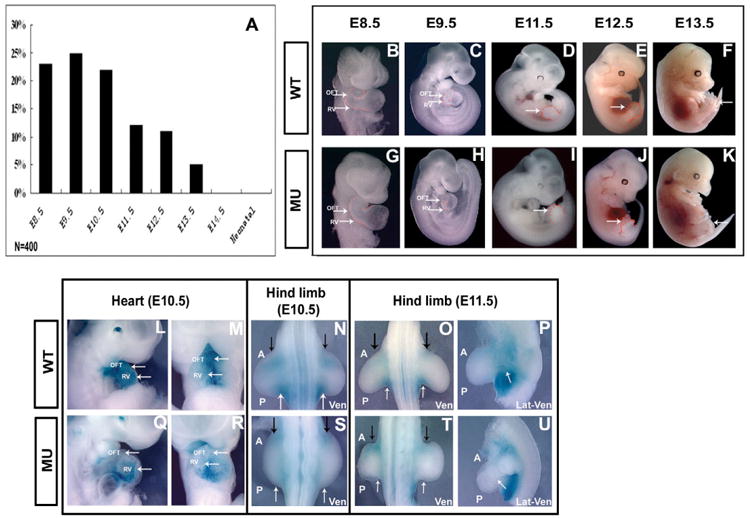
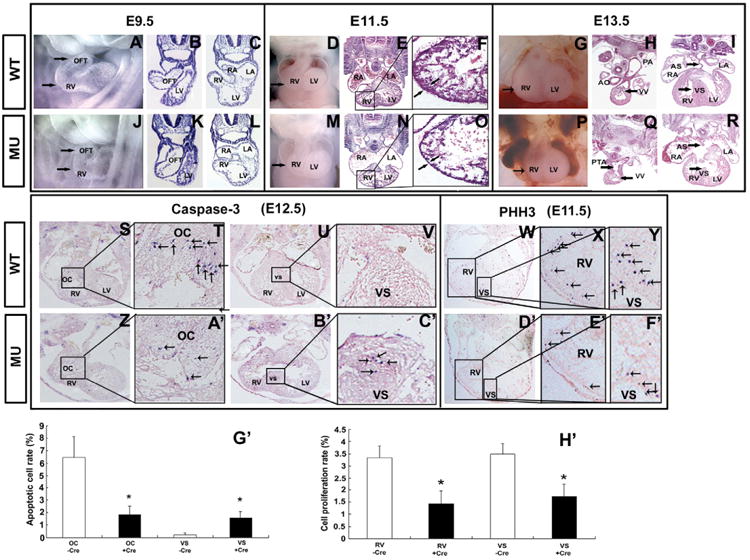
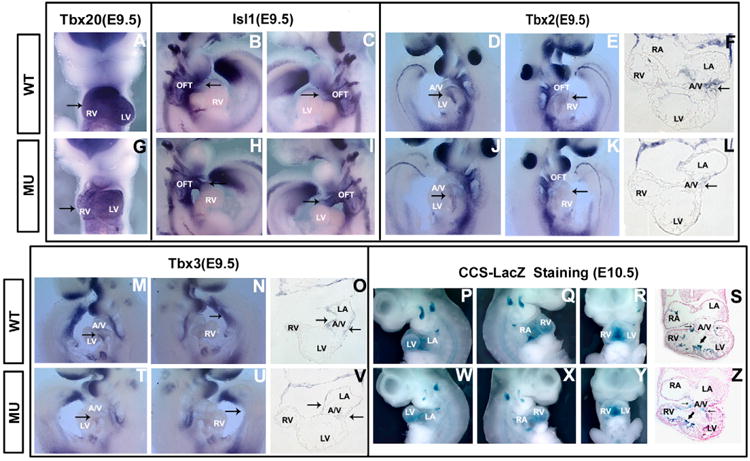
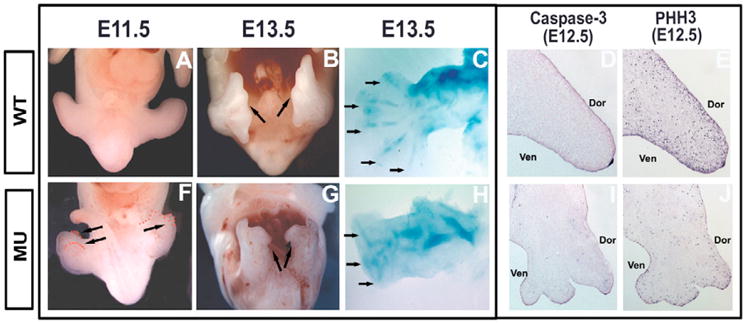
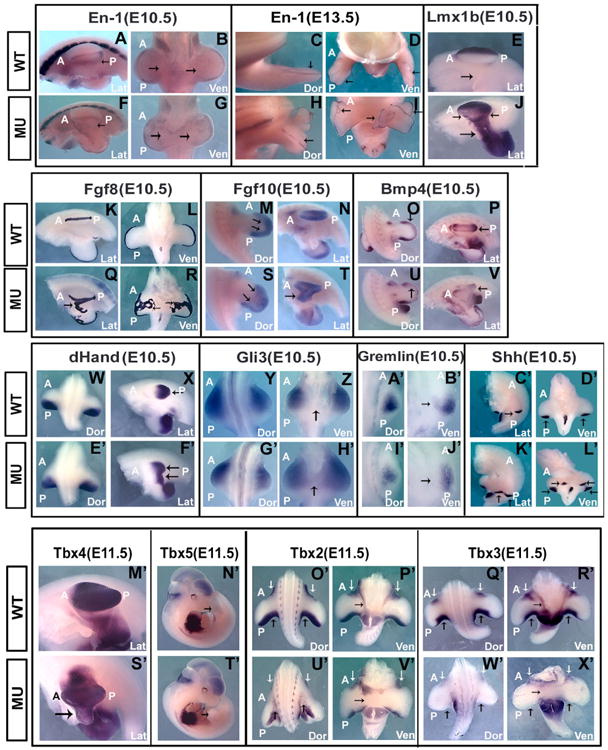
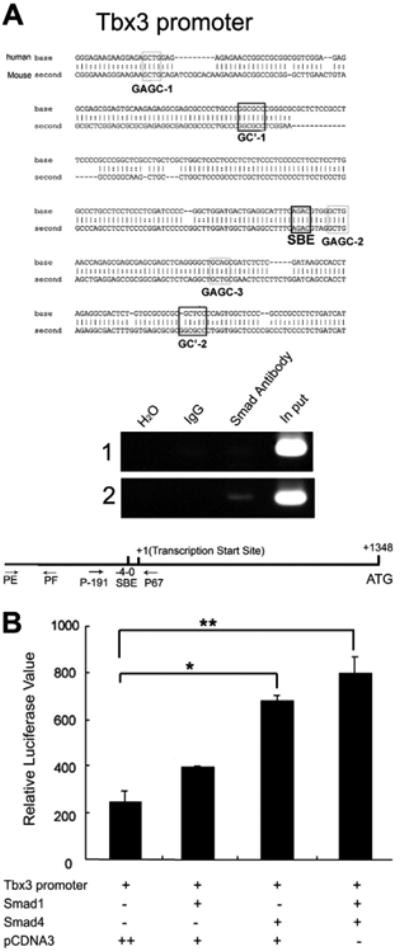
A number of human congenital disorders present with both heart and limb defects, consistent with common genetic pathways. We have recently shown that the LIM homeodomain transcription factor islet 1 (Isl1) marks a subset of cardiac progenitors. Here, we perform lineage studies with an Isl1Cre mouse line to demonstrate that Isl1 also marks a subset of limb progenitors. In both cardiac and limb progenitors, Isl1 expression is downregulated as progenitors migrate in to form either heart or limb. To investigate common heart-limb pathways in Isl1-expressing progenitors, we ablated the Type I Bmp receptor, Bmpr1a utilizing Isl1Cre/+. Analysis of consequent heart and limb phenotypes has revealed novel requirements for Bmp signaling. Additionally, we find that Bmp signaling in Isl1-expressing progenitors is required for expression of T-box transcription factors Tbx2 and Tbx3 in heart and limb. Tbx3 is required for heart and limb formation, and is mutated in ulnar-mammary syndrome. We provide evidence that the Tbx3 promoter is directly regulated by Bmp Smads in vivo.
Figures
References
-
- Ahn K, Mishina Y, Hanks MC, Behringer RR, Crenshaw EB., 3rd BMPR-IA signaling is required for the formation of the apical ectodermal ridge and dorsal-ventral patterning of the limb. Development. 2001;128:4449–4461. - PubMed
-
- Ahn S, Joyner AL. Dynamic changes in the response of cells to positive hedgehog signaling during mouse limb patterning. Cell. 2004;118:505–516. - PubMed
-
- Bardot B, Lecoin L, Fliniaux I, Huillard E, Marx M, Viallet JP. Drm/Gremlin, a BMP antagonist, defines the interbud region during feather development. Int J Dev Biol. 2004;48:149–156. - PubMed
-
- Barr M, Jr, Blackburn WR, Cooley NR., Jr Human fetal somatic and visceral morphometrics. Teratology. 1994;49:487–496. - PubMed
-
- Basson CT, Bachinsky DR, Lin RC, Levi T, Elkins JA, Soults J, Grayzel D, Kroumpouzou E, Traill TA, Leblanc-Straceski J, et al. Mutations in human TBX5 [corrected] cause limb and cardiac malformation in Holt-Oram syndrome. Nat Genet. 1997;15:30–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
