

Processing of human cathepsin D is independent of its catalytic function and auto-activation: involvement of cathepsins L and B
- PMID: 16567401
- PMCID: PMC2376303
- DOI: 10.1093/jb/mvj037
Processing of human cathepsin D is independent of its catalytic function and auto-activation: involvement of cathepsins L and B
Abstract
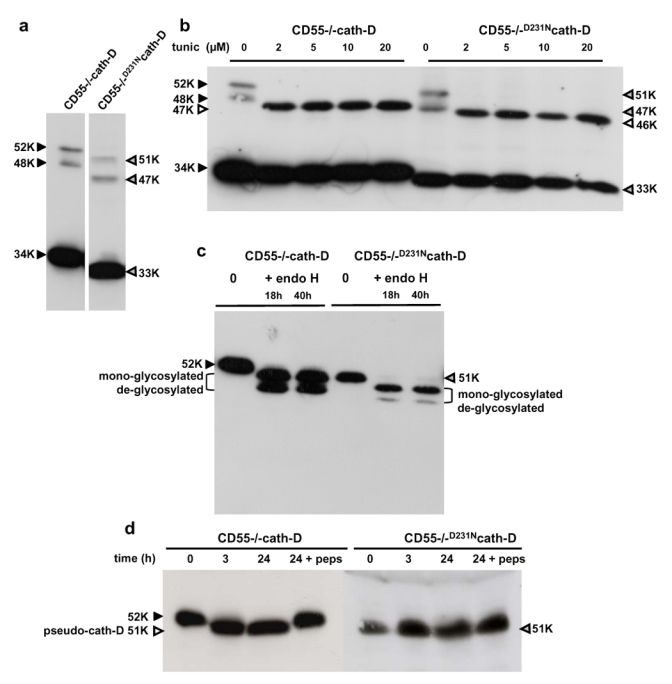
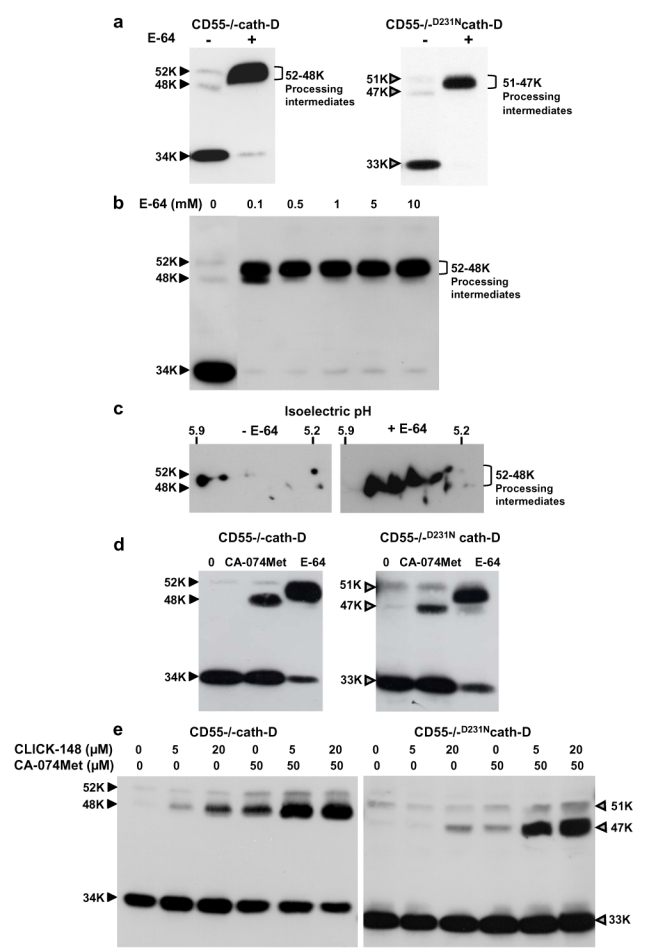
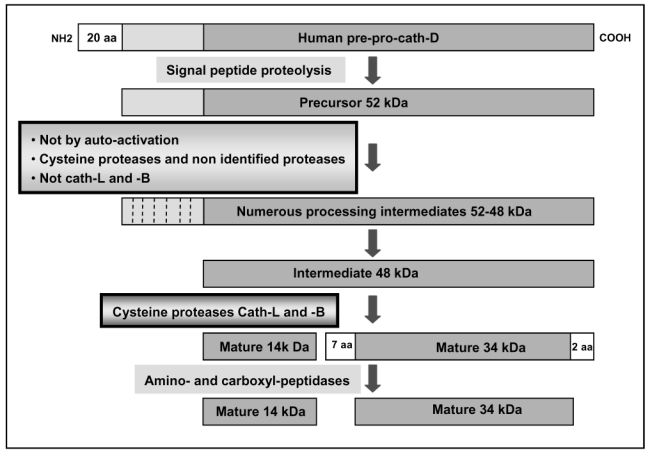
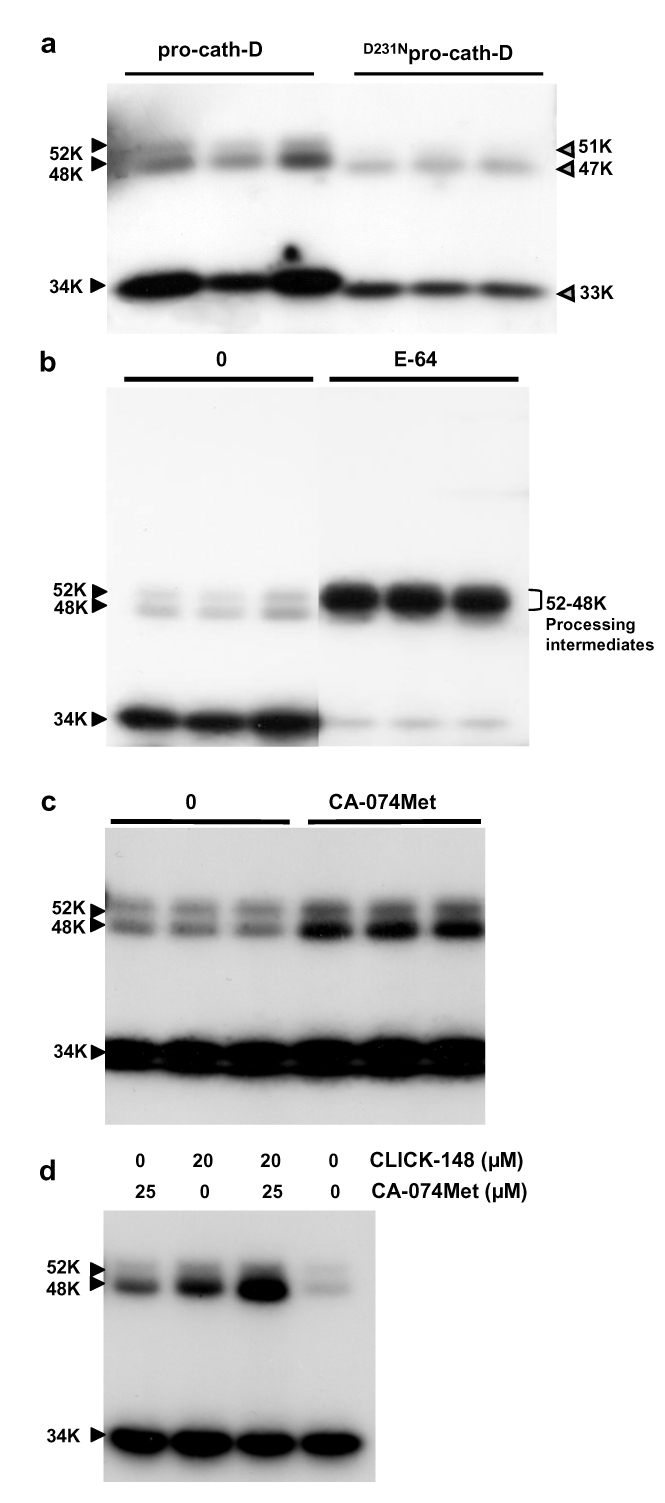
The current mechanism proposed for the processing and activation of the 52 kDa lysosomal aspartic protease cathepsin D (cath-D) is a combination of partial auto-activation generating a 51 kDa pseudo-cath-D, followed by enzyme-assisted maturation involving cysteine and/or aspartic proteases and yielding successively a 48 kDa intermediate and then 34 + 14 kDa cath-D mature species. Here we have investigated the in vivo processing of human cath-D in a cath-D-deficient fibroblast cell line in order to determine whether its maturation occurs through already active cath-D and/or other proteases. We demonstrate that cellular cath-D is processed in a manner independent of its catalytic function and that auto-activation is not a required step. Moreover, the cysteine protease inhibitor E-64 partially blocks processing, leading to accumulation of 52-48 kDa cath-D intermediates. Furthermore, two inhibitors, CLICK148 and CA-074Met, specific for the lysosomal cath-L and cath-B cysteine proteases induce accumulation of 48 kDa intermediate cath-D. Finally, maturation of endocytosed pro-cath-D is also independent of its catalytic function and requires cysteine proteases. We therefore conclude that the mechanism of cath-D maturation involves a fully-assisted processing similar to that of pro-renin.
Figures
Similar articles
-
Human recombinant pro-dipeptidyl peptidase I (cathepsin C) can be activated by cathepsins L and S but not by autocatalytic processing.Biochemistry. 2001 Feb 13;40(6):1671-8. doi: 10.1021/bi001693z. Biochemistry. 2001. PMID: 11327826
-
A role for cathepsin E in the processing of mast-cell carboxypeptidase A.J Cell Sci. 2005 May 1;118(Pt 9):2035-42. doi: 10.1242/jcs.02333. J Cell Sci. 2005. PMID: 15860733
-
Processing of cathepsins L, B and D in psoriatic epidermis.Arch Dermatol Res. 1997 Jan;289(2):87-93. doi: 10.1007/s004030050160. Arch Dermatol Res. 1997. PMID: 9049042
-
Towards specific functions of lysosomal cysteine peptidases: phenotypes of mice deficient for cathepsin B or cathepsin L.Biol Chem. 2001 May;382(5):735-41. doi: 10.1515/BC.2001.089. Biol Chem. 2001. PMID: 11517926 Review.
-
The role of thiol proteases in tissue injury and remodeling.Am J Respir Crit Care Med. 1994 Dec;150(6 Pt 2):S155-9. doi: 10.1164/ajrccm/150.6_Pt_2.S155. Am J Respir Crit Care Med. 1994. PMID: 7952652 Review.
Cited by
-
Cystatins in immune system.J Cancer. 2013;4(1):45-56. doi: 10.7150/jca.5044. Epub 2012 Dec 20. J Cancer. 2013. PMID: 23386904 Free PMC article.
-
Progranulin Stimulates the In Vitro Maturation of Pro-Cathepsin D at Acidic pH.J Mol Biol. 2019 Mar 1;431(5):1038-1047. doi: 10.1016/j.jmb.2019.01.027. Epub 2019 Jan 25. J Mol Biol. 2019. PMID: 30690031 Free PMC article.
-
Cathepsin D inhibitors as potential therapeutics for breast cancer treatment: Molecular docking and bioevaluation against triple-negative and triple-positive breast cancers.Mol Divers. 2016 May;20(2):521-35. doi: 10.1007/s11030-015-9645-8. Epub 2015 Nov 13. Mol Divers. 2016. PMID: 26563150
-
Mahogunin regulates fusion between amphisomes/MVBs and lysosomes via ubiquitination of TSG101.Cell Death Dis. 2015 Nov 5;6(11):e1970. doi: 10.1038/cddis.2015.257. Cell Death Dis. 2015. PMID: 26539917 Free PMC article.
-
Mechanisms regulating the sorting of soluble lysosomal proteins.Biosci Rep. 2022 May 27;42(5):BSR20211856. doi: 10.1042/BSR20211856. Biosci Rep. 2022. PMID: 35394021 Free PMC article. Review.
References
-
- Tang J, Wong RN. Evolution in the structure and function of aspartic proteases. J Cell Biochem. 1987;33:53–63. - PubMed
-
- Hsueh WA, Baxter JD. Human prorenin. Hypertension. 1991;17:469–477. - PubMed
-
- Richo G, Conner GE. Proteolytic activation of human procathepsin D. Adv Exp Med Biol. 1991;306:289–296. - PubMed
-
- Conner GE, Richo G. Isolation and characterization of a stable activation intermediate of the lysosomal aspartyl protease cathepsin D. Biochem. 1992;31:1142–1147. - PubMed
-
- Larsen LB, Boisen A, Petersen TE. Procathepsin D cannot autoactivate to cathepsin D at acid pH. FEBS Lett. 1993;319:54–58. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
