

A functional AMPA receptor-calcium channel complex in the postsynaptic membrane
- PMID: 16567654
- PMCID: PMC1459393
- DOI: 10.1073/pnas.0601289103
A functional AMPA receptor-calcium channel complex in the postsynaptic membrane
Abstract
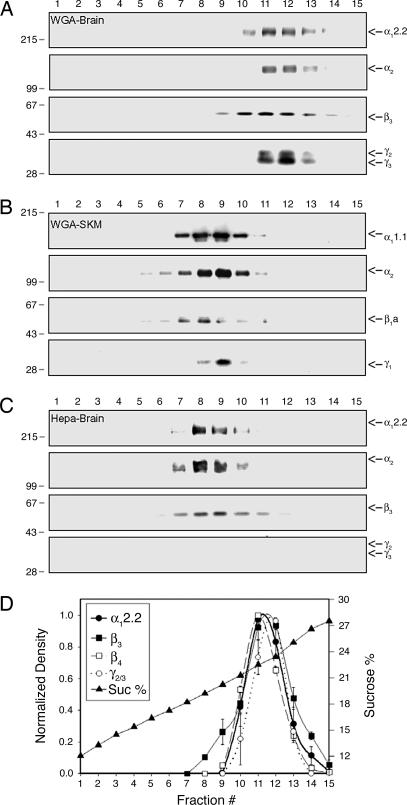
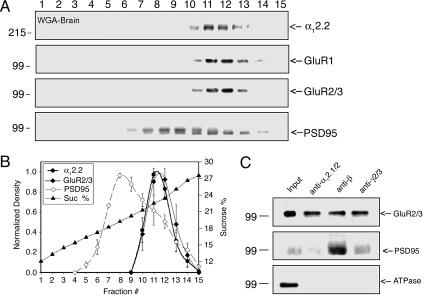
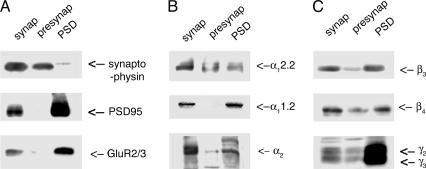
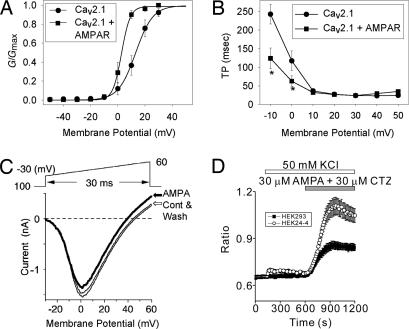
Ca(2+) channels play critical roles in the regulation of synaptic activity. In contrast to the well established function of voltage-activated Ca(2+) channels in the presynaptic membrane for neurotransmitter release, some studies are just beginning to elucidate the functions of the Ca(2+) channels in the postsynaptic membrane. In this study, we demonstrated the functional association of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptors with the neuronal Ca(2+) channels. A series of biochemical studies showed the specific association of Ca(v)2.1 (alpha(1A)-class) and Ca(v)2.2 (alpha(1B)-class) with AMPA receptors in the postsynaptic membrane. Our electrophysiological and Ca(2+) imaging analyses of recombinant Ca(v)2.1 and AMPA receptors also showed functional coupling of the two channels. Considering the critical roles of postsynaptic intracellular concentration of Ca(2+) ([Ca(2+)](i)) increase and AMPA receptor trafficking for long-term potentiation (LTP) and long-term depression (LTD), the functional association of Ca(2+) channels with the AMPA receptors may provide new insights into the mechanism of synaptic plasticity.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Similar articles
-
Loss of calcium channels in the cerebellum of the ataxic and epileptic stargazer mutant mouse.Brain Res. 2009 Jul 7;1279:156-67. doi: 10.1016/j.brainres.2009.04.051. Epub 2009 May 5. Brain Res. 2009. PMID: 19422811
-
Input- and subunit-specific AMPA receptor trafficking underlying long-term potentiation at hippocampal CA3 synapses.Eur J Neurosci. 2004 Jul;20(1):101-10. doi: 10.1111/j.1460-9568.2004.03461.x. Eur J Neurosci. 2004. PMID: 15245483
-
AMPA induced Ca2+ influx in motor neurons occurs through voltage gated Ca2+ channel and Ca2+ permeable AMPA receptor.Neurochem Int. 2011 Nov;59(6):913-21. doi: 10.1016/j.neuint.2011.06.023. Epub 2011 Jul 13. Neurochem Int. 2011. PMID: 21777635
-
Functional modulation of AMPA receptors by transmembrane AMPA receptor regulatory proteins.Neuroscience. 2009 Jan 12;158(1):45-54. doi: 10.1016/j.neuroscience.2007.12.046. Epub 2008 Jan 18. Neuroscience. 2009. PMID: 18304745 Review.
-
Neurophysiology of HCN channels: from cellular functions to multiple regulations.Prog Neurobiol. 2014 Jan;112:1-23. doi: 10.1016/j.pneurobio.2013.10.001. Epub 2013 Oct 29. Prog Neurobiol. 2014. PMID: 24184323 Review.
Cited by
-
Stargazin-related protein γ₇ is associated with signalling endosomes in superior cervical ganglion neurons and modulates neurite outgrowth.J Cell Sci. 2011 Jun 15;124(Pt 12):2049-57. doi: 10.1242/jcs.084988. Epub 2011 May 24. J Cell Sci. 2011. PMID: 21610096 Free PMC article.
-
Glutamate-induced calcium transients in rat neurons of the dorsal motor nucleus of the vagus.J Gastrointest Surg. 2007 Aug;11(8):1016-24. doi: 10.1007/s11605-007-0176-1. J Gastrointest Surg. 2007. PMID: 17549578
-
Rab3-interacting molecule gamma isoforms lacking the Rab3-binding domain induce long lasting currents but block neurotransmitter vesicle anchoring in voltage-dependent P/Q-type Ca2+ channels.J Biol Chem. 2010 Jul 9;285(28):21750-67. doi: 10.1074/jbc.M110.101311. Epub 2010 May 7. J Biol Chem. 2010. PMID: 20452978 Free PMC article.
-
Reactive oxygen species (ROS) modulate AMPA receptor phosphorylation and cell-surface localization in concert with pain-related behavior.Pain. 2012 Sep;153(9):1905-1915. doi: 10.1016/j.pain.2012.06.001. Epub 2012 Jul 4. Pain. 2012. PMID: 22770842 Free PMC article.
-
Supramolecular assemblies and localized regulation of voltage-gated ion channels.Physiol Rev. 2009 Apr;89(2):411-52. doi: 10.1152/physrev.00029.2007. Physiol Rev. 2009. PMID: 19342611 Free PMC article. Review.
References
-
- Song I., Huganir R. L. Trends Neurosci. 2002;25:578–588. - PubMed
-
- Kovalchuk Y., Hanse E., Kafitz K. W., Konnerth A. Science. 2002;295:1729–1734. - PubMed
-
- Witcher D. R., De Waard M., Kahl S.D., Campbell K.P. Methods Enzymol. 1994;238:335–348. - PubMed
-
- Kang M. G., Chen C. C., Felix R., Letts V. A., Frankel W. N., Mori Y., Campbell K. P. J. Biol. Chem. 2001;276:32917–32924. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
