

Functional comparison of mouse CIRE/mouse DC-SIGN and human DC-SIGN
- PMID: 16569675
- PMCID: PMC7185610
- DOI: 10.1093/intimm/dxl011
Functional comparison of mouse CIRE/mouse DC-SIGN and human DC-SIGN
Abstract
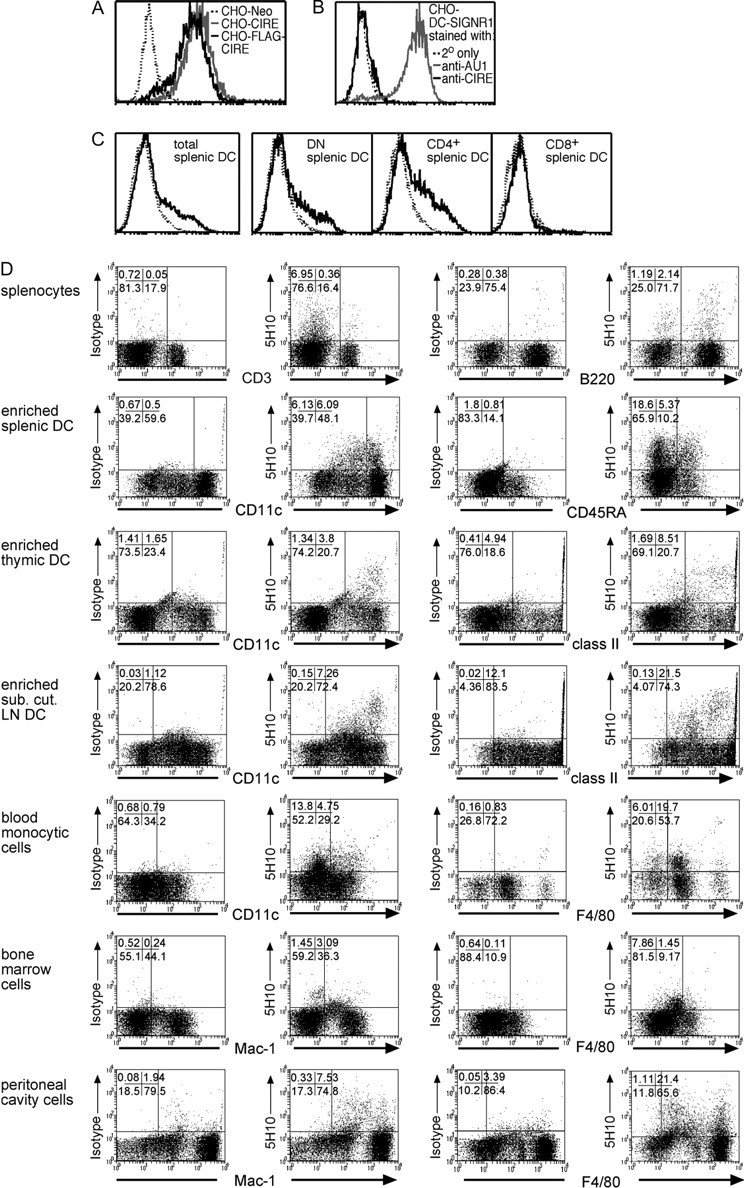
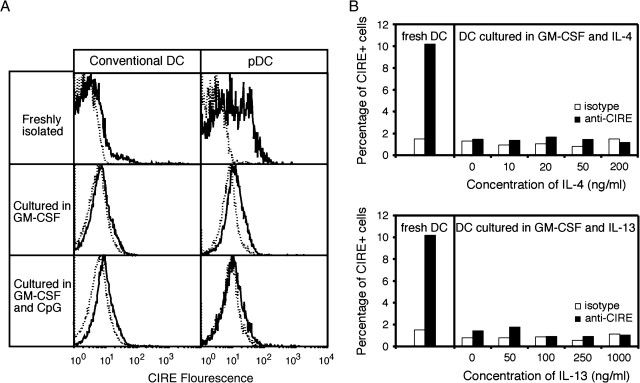
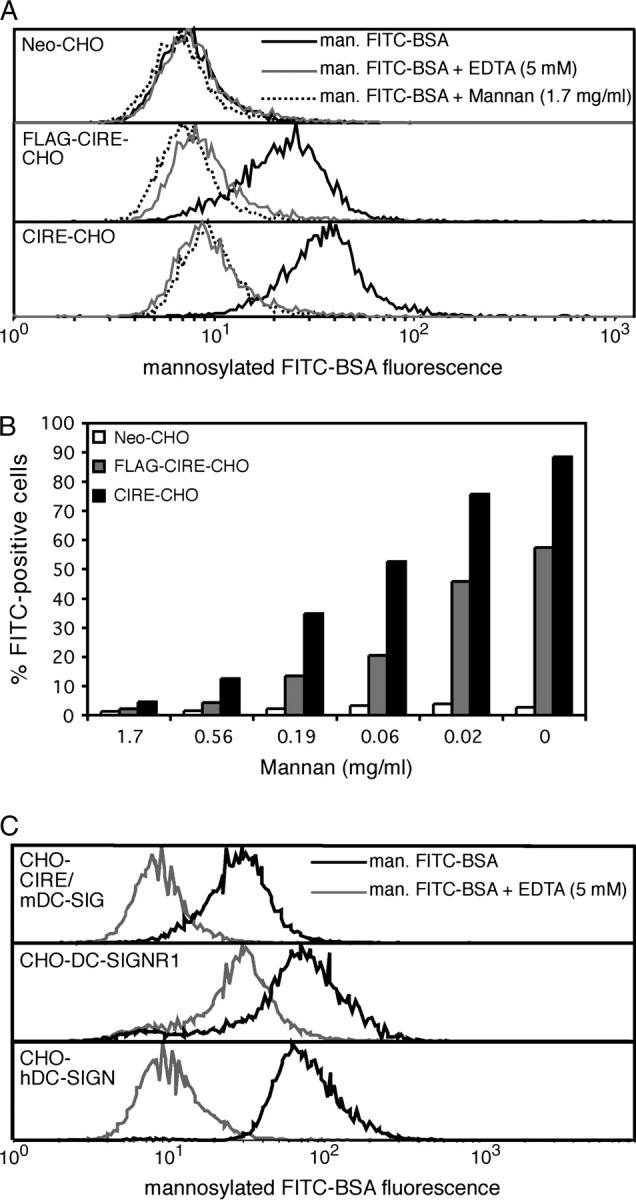
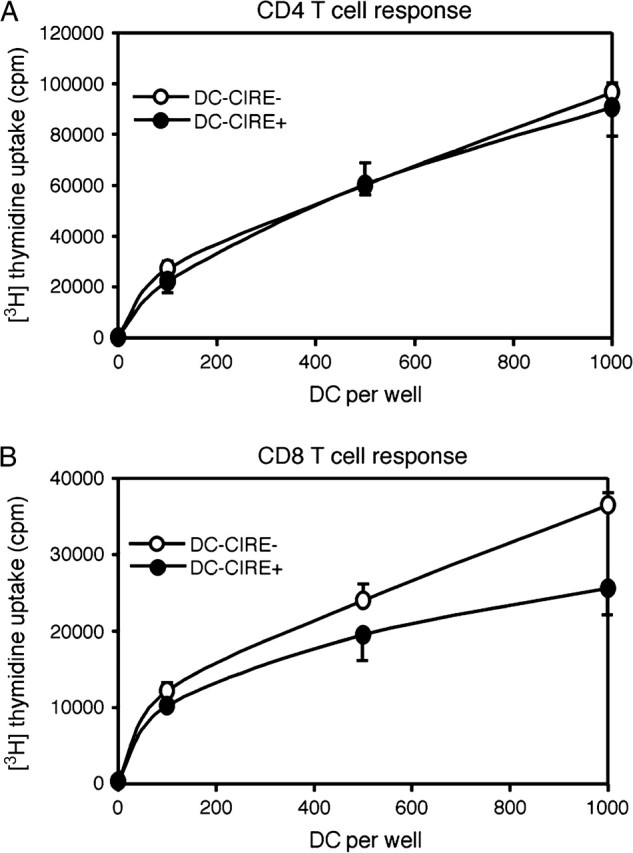
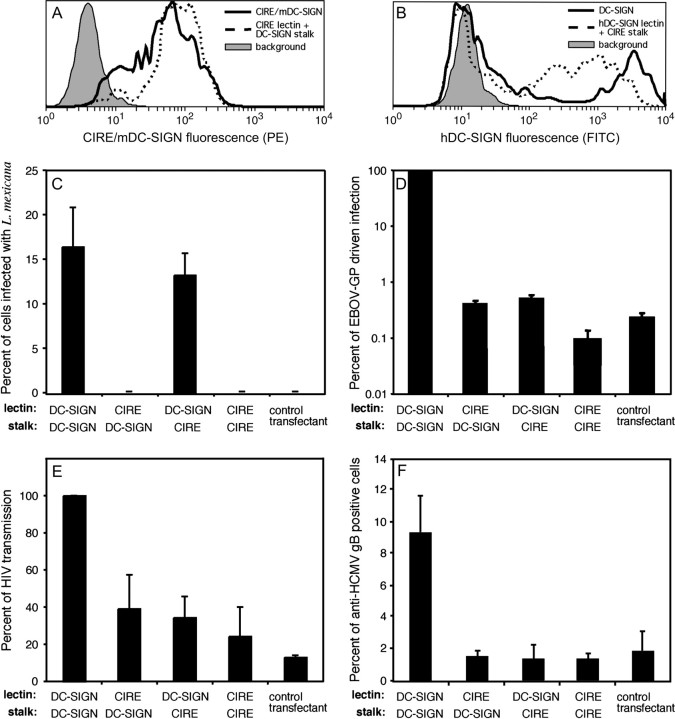
CIRE/mDC-SIGN is a C-type lectin we originally identified as a molecule differentially expressed by mouse dendritic cell (DC) populations. Immunostaining with a CIRE/mDC-SIGN-specific mAb revealed that CIRE/mDC-SIGN is indeed on the surface of some CD4+, CD4- 8- DCs and plasmacytoid pre-DCs, but not on CD8+ DCs. It has been proposed that CIRE/mDC-SIGN is the functional orthologue of human DC-SIGN (hDC-SIGN), a molecule that both enhances T cell responses and facilitates antigen uptake. We assessed if CIRE/mDC-SIGN and hDC-SIGN exhibit functional similarities. CIRE/mDC-SIGN is down-regulated upon activation, but unlike hDC-SIGN, incubation with IL-4 and IL-13 did not enhance CIRE/mDC-SIGN expression, indicating differences in gene regulation. Like hDC-SIGN, CIRE/mDC-SIGN bound mannosylated residues. However, we could detect no role for CIRE/mDC-SIGN in T cell-DC interactions and the protein did not bind to pathogens known to interact with hDC-SIGN, including Leishmania mexicana, cytomegalovirus, HIV and lentiviral particles bearing the Ebolavirus glycoprotein. The binding of CIRE/mDC-SIGN to hDC-SIGN ligands was not rescued when CIRE/mDC-SIGN was engineered to express the stalk region of hDC-SIGN. We conclude that there are significant differences in the fine specificity of the C-type lectin domains of hDC-SIGN and CIRE/mDC-SIGN and that these two molecules may not be functional orthologues.
Figures

References
-
- Steinman R. M. 1991. The dendritic cell system and its role in immunogenicity. Annu. Rev. Immunol. 9:271. - PubMed
-
- Shortman K. and Liu Y. J. 2002. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2:151. - PubMed
-
- Vremec D., Pooley J., Hochrein H., Wu L., and Shortman K. 2000. CD4 and CD8 expression by dendritic cell subtypes in mouse thymus and spleen. J. Immunol. 164:2978. - PubMed
-
- Shortman K. and Wu L. 2001. Parentage and heritage of dendritic cells. Blood 97:3325. - PubMed
-
- Traver D., Akashi K., Manz M. et al.2000. Development of CD8alpha-positive dendritic cells from a common myeloid progenitor. Science 290:2152. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
