

Inactivating a cellular intrinsic immune defense mediated by Daxx is the mechanism through which the human cytomegalovirus pp71 protein stimulates viral immediate-early gene expression
- PMID: 16571803
- PMCID: PMC1440479
- DOI: 10.1128/JVI.80.8.3863-3871.2006
Inactivating a cellular intrinsic immune defense mediated by Daxx is the mechanism through which the human cytomegalovirus pp71 protein stimulates viral immediate-early gene expression
Abstract
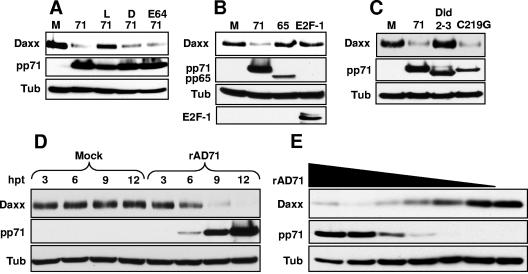
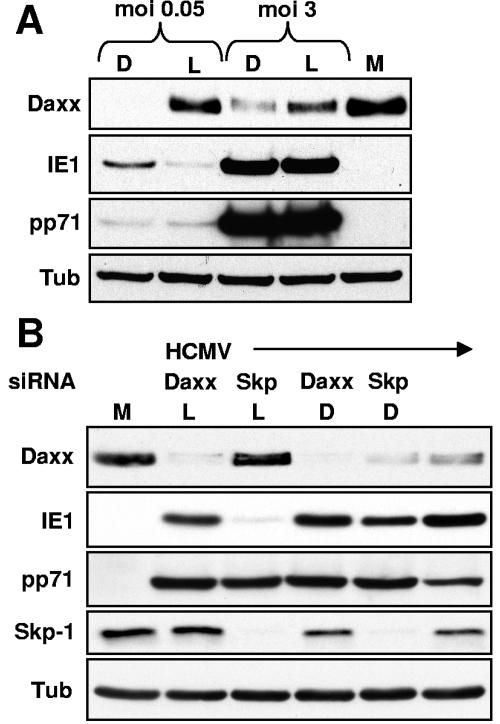
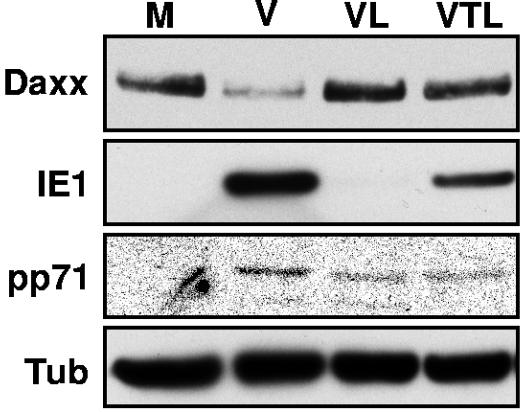
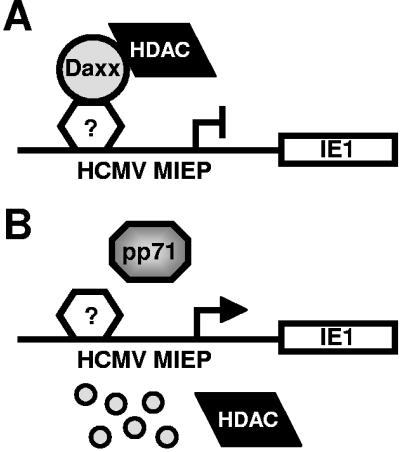
Human cytomegalovirus (HCMV) masterfully evades adaptive and innate immune responses, allowing infection to be maintained and periodically reactivated for the life of the host. Here we show that cells also possess an intrinsic immune defense against HCMV that is disarmed by the virus. In HCMV-infected cells, the promyelocytic leukemia nuclear body (PML-NB) protein Daxx silences viral immediate-early gene expression through the action of a histone deacetylase. However, this antiviral tactic is efficiently neutralized by the viral pp71 protein, which is incorporated into virions, delivered to cells upon infection, and mediates the proteasomal degradation of Daxx. This work demonstrates the mechanism through which pp71 activates viral immediate-early gene expression in HCMV-infected cells. Furthermore, it provides insight into how a PML-NB protein institutes an intrinsic immune defense against a DNA virus and how HCMV pp71 inactivates this defense.
Figures
Similar articles
-
Human cytomegalovirus gene expression is silenced by Daxx-mediated intrinsic immune defense in model latent infections established in vitro.J Virol. 2007 Sep;81(17):9109-20. doi: 10.1128/JVI.00827-07. Epub 2007 Jun 27. J Virol. 2007. PMID: 17596307 Free PMC article.
-
Proteasome-dependent, ubiquitin-independent degradation of Daxx by the viral pp71 protein in human cytomegalovirus-infected cells.Virology. 2007 Oct 25;367(2):334-8. doi: 10.1016/j.virol.2007.05.037. Epub 2007 Jun 27. Virology. 2007. PMID: 17590404
-
Daxx-mediated accumulation of human cytomegalovirus tegument protein pp71 at ND10 facilitates initiation of viral infection at these nuclear domains.J Virol. 2002 Aug;76(15):7705-12. doi: 10.1128/jvi.76.15.7705-7712.2002. J Virol. 2002. PMID: 12097584 Free PMC article.
-
Immediate-Early (IE) gene regulation of cytomegalovirus: IE1- and pp71-mediated viral strategies against cellular defenses.Virol Sin. 2014 Dec;29(6):343-52. doi: 10.1007/s12250-014-3532-9. Epub 2014 Nov 25. Virol Sin. 2014. PMID: 25501994 Free PMC article. Review.
-
Intrinsic cellular defense mechanisms targeting human cytomegalovirus.Virus Res. 2011 May;157(2):128-33. doi: 10.1016/j.virusres.2010.10.002. Epub 2010 Oct 8. Virus Res. 2011. PMID: 20934469 Review.
Cited by
-
The Role of Nuclear Antiviral Factors against Invading DNA Viruses: The Immediate Fate of Incoming Viral Genomes.Viruses. 2016 Oct 22;8(10):290. doi: 10.3390/v8100290. Viruses. 2016. PMID: 27782081 Free PMC article. Review.
-
Epigenetics and Genetics of Viral Latency.Cell Host Microbe. 2016 May 11;19(5):619-28. doi: 10.1016/j.chom.2016.04.008. Cell Host Microbe. 2016. PMID: 27173930 Free PMC article. Review.
-
Human Cytomegalovirus Latency: Approaching the Gordian Knot.Annu Rev Virol. 2016 Sep 29;3(1):333-357. doi: 10.1146/annurev-virology-110615-042422. Epub 2016 Aug 4. Annu Rev Virol. 2016. PMID: 27501258 Free PMC article.
-
The Abundant Tegument Protein pUL25 of Human Cytomegalovirus Prevents Proteasomal Degradation of pUL26 and Supports Its Suppression of ISGylation.J Virol. 2018 Nov 27;92(24):e01180-18. doi: 10.1128/JVI.01180-18. Print 2018 Dec 15. J Virol. 2018. PMID: 30282718 Free PMC article.
-
Cell Line Models for Human Cytomegalovirus Latency Faithfully Mimic Viral Entry by Macropinocytosis and Endocytosis.J Virol. 2019 Oct 15;93(21):e01021-19. doi: 10.1128/JVI.01021-19. Print 2019 Nov 1. J Virol. 2019. PMID: 31391271 Free PMC article.
References
-
- Arnon, T. I., H. Achdout, O. Levi, G. Markel, N. Saleh, G. Katz, R. Gazit, T. Gonen-Gross, J. Hanna, E. Nahari, A. Porgador, A. Honigman, B. Plachter, D. Mevorach, D. G. Wolf, and O. Mandelboim. 2005. Inhibition of the NKp30 activating receptor by pp65 of human cytomegalovirus. Nat. Immunol. 6:515-523. - PubMed
-
- Bieniasz, P. D. 2004. Intrinsic immunity: a front-line defense against viral attack. Nat. Immunol. 5:1109-1115. - PubMed
-
- Brooks, A. R., R. N. Harkins, P. Wang, H. S. Qian, P. Liu, and G. M. Rubanyi. 2004. Transcriptional silencing is associated with extensive methylation of the CMV promoter following adenoviral gene delivery to muscle. J. Gene Med. 6:395-404. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
