

doi: 10.1128/JVI.80.8.4168-4173.2006.
Expression of baculovirus late and very late genes depends on LEF-4, a component of the viral RNA polymerase whose guanyltransferase function is essential
Affiliations
- PMID: 16571832
- PMCID: PMC1440449
- DOI: 10.1128/JVI.80.8.4168-4173.2006
Item in Clipboard
Expression of baculovirus late and very late genes depends on LEF-4, a component of the viral RNA polymerase whose guanyltransferase function is essential
J Virol.
2006 Apr.
Abstract
Baculovirus lef-4 encodes one subunit of the viral RNA polymerase. Here, we demonstrate the essential nature of LEF-4 by RNA interference and bacmid knockout technology. Silencing of LEF-4 in wild-type virus-infected cells suppressed expression of structural genes, while early expression was unaffected, demonstrating its essential role in late gene expression. After transfection of insect cells with lef-4 mutant bacmid, no viral progeny was produced, further defining its central role in infection. Cotransfection with wild-type lef-4 plasmid restored normal replication, but plasmid encoding a guanyltransferase-deficient version failed to rescue. These results emphasize the importance of the mRNA capping function of LEF-4.
Figures
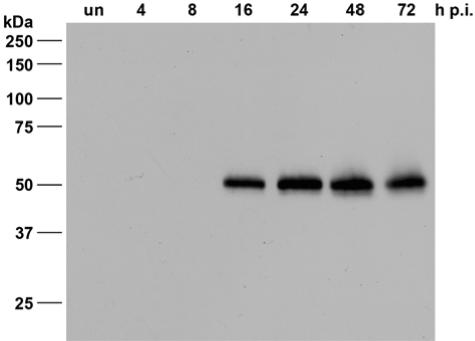
Expression of LEF-4 in AcMNPV-infected S. frugiperda cells. Detergent-based nuclear extracts were prepared from uninfected S. frugiperda cells (“un”) or from infected cells at 4, 8, 16, 24, 48, and 72 h p.i. Proteins were resolved on sodium dodecyl sulfate-10% polyacrylamide gels and stained with the rabbit anti-LEF-4 antiserum. Protein size markers are given on the left.
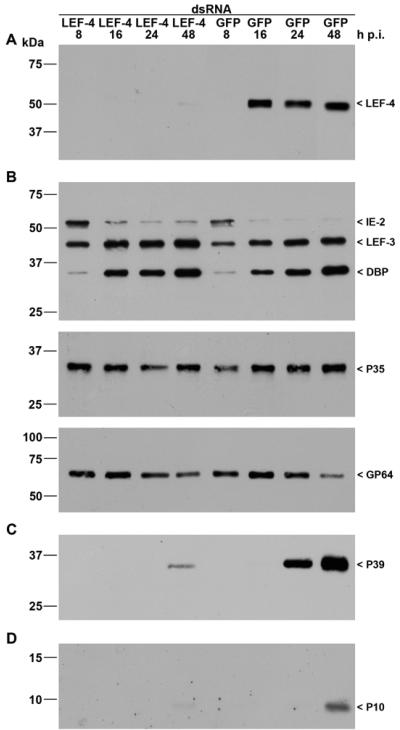
Inhibition of viral gene expression by lef-4 silencing. S. frugiperda cells were transfected with either LEF-4 or GFP dsRNA. Cells were subsequently infected with AcMNPV (10 PFU/cell) at 20 h posttransfection, and detergent-based nuclear and cytoplasmic extracts were prepared from infected cells 8, 16, 24, and 48 h p.i. Proteins were resolved on sodium dodecyl sulfate-10% polyacrylamide gels and transferred to nitrocellulose membranes (A, B, and C) or resolved on sodium dodecyl sulfate-15% polyacrylamide gels and transferred to polyvinylidene difluoride membranes (D). (A) LEF-4 was stained with rabbit anti-Lef-4 antiserum. (B) Early gene expression was analyzed with rabbit antisera raised against IE2 (16), LEF-3 (3), DBP (22), or P35 (11) or with mouse monoclonal anti-GP64 (AcV5) (12). (C) Late gene expression was analyzed with mouse monoclonal anti-P39 (P10C6) (30). (D) Expression of the very late protein P10 was detectable with rabbit anti-P10 serum (29). Expression of LEF-4, IE2, LEF-3, DBP, P39, and P10 was analyzed on samples of nuclear protein fractions, and P35 and GP64 expression was detected in cytoplasmic fractions as described previously (21). Protein size markers are shown on the left, and the identities of the viral proteins are indicated on the right.
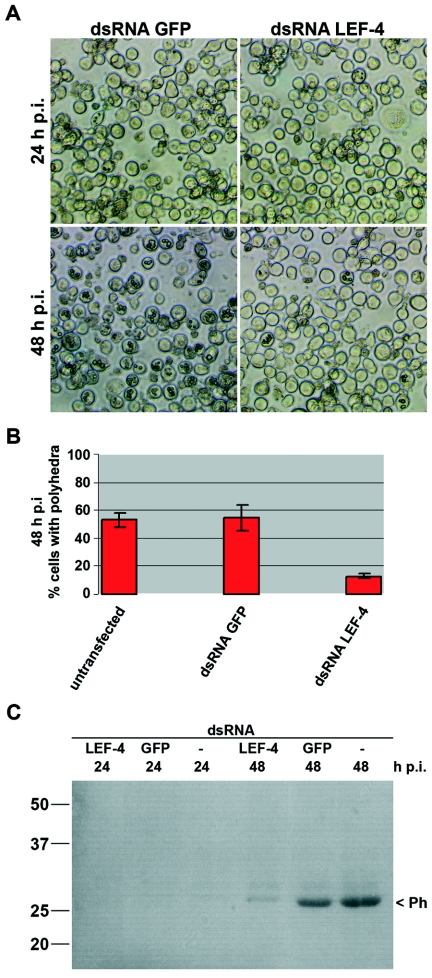
Polyhedron formation and polyhedrin expression upon lef-4 inhibition in AcMNPV-infected S. frugiperda cells. Cells were transfected with either LEF-4 dsRNA or GFP dsRNA, infected with AcMNPV (10 PFU/cells) at 20 h posttransfection, and analyzed at 24 and 48 h p.i. (A) Phase-contrast images are shown, and (B) polyhedron-containing cells were quantitated at 48 h p.i. (C) Cells transfected with LEF-4 dsRNA, GFP dsRNA, or untransfected cells were infected, and detergent-based nuclear extracts were prepared at 24 and 48 h p.i. Proteins were resolved on sodium dodecyl sulfate-10% polyacrylamide gels and transferred to nitrocellulose, and polyhedrin was viewed by Ponceau staining. Numbers at left are molecular masses in kilodaltons.
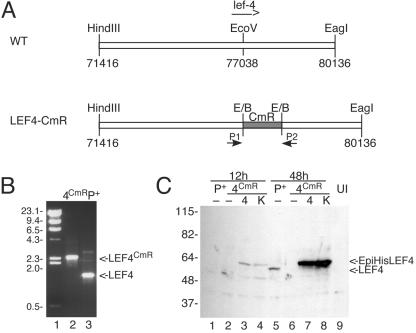
Construction of BacP+/LEF-4CmR. (A) Cloning strategy. A plasmid containing the left 7 kb of the HindIII-C fragment of AcMNPV was digested with EcoRV, and a 1-kb fragment of pBR325 containing the CAT resistance marker was inserted. The resultant plasmid was transformed into BJ5183 cells containing a modified version of the AcMNPV genome, followed by selection on chloramphenicol, as previously described (1). (B) PCR screening. Correct insertion of CAT into lef-4 was verified by PCR using primers that flank the lef-4 open reading frame. Lane 2, BacP+/LEF-4CmR; lane 3, BacP+. The positions of molecular mass markers (lane 1) are indicated on the left in kilodaltons, and the migration of lef-4 and lef-4 with the CAT insertion is shown on the right. (C) Immunoblot analysis of LEF-4 expression in transfected cells. Cells were transfected with BacP+ (P+, lanes 1 and 5) or BacP+/LEF-4CmR (4CmR, lanes 2 to 4 and 6 to 8) and cotransfected where indicated with pHSEpiHisLEF-4 (4, lanes 3 and 7) or pHSEpiHisLEF-4(K255A) (K, lanes 4 and 8). Cells were harvested at 12 h (lanes 1 to 4) or 48 h (lanes 5 to 8) posttransfection. The blot was probed with rabbit LEF-4 antiserum. Untransfected cells were analyzed as a negative control (lane 9). The positions of molecular mass markers are indicated on the left in kilodaltons, and the migration of LEF-4 and EpiHisLEF-4 is indicated on the right.

Infectivity of lef-4 mutant viruses. S. frugiperda cells were transfected with BacP+ (A and E), BacP+/LEF-4CmR (B and F), or BacP+/LEF-4CmR in the presence of plasmids encoding wild-type LEF-4 (C and G) or a mutant version of LEF-4 that lacks guanyltransferase activity (D and H). Cells were harvested at 12 or 48 h p.i. and processed for immunofluorescence using a mouse monoclonal antibody raised against IE1. Images were collected using a Zeiss ApoTome/Axioplan 2 microscope. Images shown represent merged images of phase contrast and green fluorescence to visualize percentages of cells expressing IE1.
Similar articles
-
Analysis of an Autographa californica nucleopolyhedrovirus lef-11 knockout: LEF-11 is essential for viral DNA replication.J Virol. 2002 Mar;76(6):2770-9. doi: 10.1128/jvi.76.6.2770-2779.2002. J Virol. 2002. PMID: 11861844 Free PMC article.
-
Functional characterization of Bombyx mori nucleopolyhedrovirus mutant lacking late expression factor 9.Acta Virol. 2016;60(3):281-9. doi: 10.4149/av_2016_03_281. Acta Virol. 2016. PMID: 27640438
-
Characterization of late gene expression factor LEF-10 from Bombyx mori nucleopolyhedrovirus.Virus Res. 2013 Jul;175(1):45-51. doi: 10.1016/j.virusres.2013.03.022. Epub 2013 Apr 17. Virus Res. 2013. PMID: 23603137
-
An Autographa californica nucleopolyhedrovirus lef-2 mutant: consequences for DNA replication and very late gene expression.Virology. 1996 Mar 1;217(1):338-48. doi: 10.1006/viro.1996.0121. Virology. 1996. PMID: 8599220
-
Deletion analysis of four of eighteen late gene expression factor gene homologues of the baculovirus, BmNPV.Virology. 1997 Mar 31;230(1):35-47. doi: 10.1006/viro.1997.8457. Virology. 1997. PMID: 9126260
Cited by
-
Promoter motifs essential to the differential transcription of structural and non-structural genes of the white spot syndrome virus.Virus Genes. 2009 Oct;39(2):223-33. doi: 10.1007/s11262-009-0380-z. Virus Genes. 2009. PMID: 19554443
-
Use of bacterial artificial chromosomes in baculovirus research and recombinant protein expression: current trends and future perspectives.ISRN Microbiol. 2012 Sep 12;2012:628797. doi: 10.5402/2012/628797. Print 2012. ISRN Microbiol. 2012. PMID: 23762754 Free PMC article.
-
Systematic Analysis of 42 Autographa Californica Multiple Nucleopolyhedrovirus Genes Identifies An Additional Six Genes Involved in the Production of Infectious Budded Virus.Virol Sin. 2021 Aug;36(4):762-773. doi: 10.1007/s12250-021-00355-1. Epub 2021 Mar 8. Virol Sin. 2021. PMID: 33683665 Free PMC article.
-
The genome of Gryllus bimaculatus nudivirus indicates an ancient diversification of baculovirus-related nonoccluded nudiviruses of insects.J Virol. 2007 May;81(10):5395-406. doi: 10.1128/JVI.02781-06. Epub 2007 Mar 14. J Virol. 2007. PMID: 17360757 Free PMC article.
-
Characterization of LEF4 ligand binding property and its role as part of baculoviral transcription machinery.Mol Cell Biochem. 2010 Jan;333(1-2):83-9. doi: 10.1007/s11010-009-0207-1. Epub 2009 Jul 25. Mol Cell Biochem. 2010. PMID: 19633819
References
-
- Bideshi, D. K., and B. A. Federici. 2000. The Trichoplusia ni granulovirus helicase is unable to support replication of Autographa californica multicapsid nucleopolyhedrovirus in cells and larvae of T. ni. J. Gen. Virol. 81:1593-1599. - PubMed
-
- Carstens, E. B., H. Chan, H. Yu, G. V. Williams, and R. Casselman. 1994. Genetic analyses of temperature-sensitive mutations in baculovirus late expression factors. Virology 204:323-337. - PubMed
-
- Flores-Jasso, C. F., V. J. Valdes, A. Sampieri, V. Valadez-Graham, F. Recillas-Targa, and L. Vaca. 2004. Silencing structural and nonstructural genes in baculovirus by RNA interference. Virus Res. 102:75-84. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
