

doi: 10.1128/JVI.80.8.4174-4178.2006.
Role of endosomal cathepsins in entry mediated by the Ebola virus glycoprotein
Affiliations
- PMID: 16571833
- PMCID: PMC1440424
- DOI: 10.1128/JVI.80.8.4174-4178.2006
Item in Clipboard
Role of endosomal cathepsins in entry mediated by the Ebola virus glycoprotein
J Virol.
2006 Apr.
Abstract
Using chemical inhibitors and small interfering RNA (siRNA), we have confirmed roles for cathepsin B (CatB) and cathepsin L (CatL) in Ebola virus glycoprotein (GP)-mediated infection. Treatment of Ebola virus GP pseudovirions with CatB and CatL converts GP1 from a 130-kDa to a 19-kDa species. Virus with 19-kDa GP1 displays significantly enhanced infection and is largely resistant to the effects of the CatB inhibitor and siRNA, but it still requires a low-pH-dependent endosomal/lysosomal function. These and other results support a model in which CatB and CatL prime GP by generating a 19-kDa intermediate that can be acted upon by an as yet unidentified endosomal/lysosomal enzyme to trigger fusion.
Figures

Effect of cathepsin inhibitors and siRNA on infection of Vero cells by VSV-GP. (A) Cells were pretreated for 2 h at 37°C with vehicle alone (V), CatB inhibitor (B), or CatL inhibitor (L) at low concentrations (0.5 μM CatB inhibitor, 1 μM CatL inhibitor) and high concentrations (1 μM CatB inhibitor, 4 μM CatL inhibitor). Cells were washed and infected with VSV-GP and VSV-G pseudovirions at approximate multiplicities of infection (MOI) of 0.2 to 0.4 and 0.5 to 1.0, respectively, in the presence of fresh inhibitors. Twenty-four hours postinfection, the cells were fixed and the percentage of GFP-positive cells was determined by flow cytometry. Results shown are the averages of normalized data from three to six experiments, where the average percent infection for vehicle-treated cells was 31 ± 19 for VSV-GP and 56 ± 10 for VSV-G. Error bars indicate standard errors, and asterisks indicate a statistically significant deviation from the mean relative to vehicle-treated cells at a confidence level of greater than 99%. (B and C) Cells were transfected with a nontargeting control siRNA oligonucleotide (Con) or siRNA oligonucleotides targeting CatB and/or CatL. Seventy-two hours posttransfection, the cells were either lysed for Western blotting and activity assays (B) or infected as described for panel A (C). (B) Representative blot of CatB and CatL expression in siRNA-treated cells and the average results from five activity assays ± the standard deviation. Results shown in panel C are the averages of normalized data from three to six experiments, where the average percent infection for control siRNA-treated cells was 23 ± 14 for VSV-GP and 53 ± 6 for VSV-G. Error bars indicate the standard errors, and asterisks indicate a statistically significant deviation from the mean relative to control siRNA-treated cells at a confidence level greater than 99%.
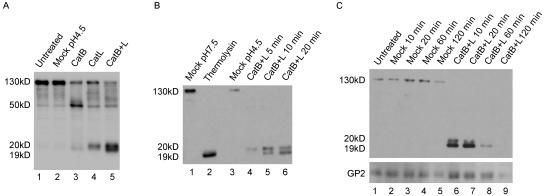
Cleavage of Ebola virus GP by CatB and CatL. (A) VSV-GP was treated with no enzyme (mock) or 20 μg/ml CatB, CatL, or CatB plus CatL (CatB+L) at pH 4.5 with 4 mM dithiothreitol for 10 min at 37°C. Reactions were quenched, and samples were analyzed by Western blotting for GP1. (B) VSV-GP was treated with 20 μg/ml CatB plus CatL in the above conditions for 5, 10, or 20 min or with 0.5 mg/ml thermolysin at pH 7.5 for 20 min. Mock treatments were performed in the same buffers containing no enzyme. Reactions were quenched and analyzed by Western blotting for GP1. (C) VSV-GP was mock treated or treated with 25 μg/ml CatB plus CatL as described above for 10, 20, 60, or 120 min. Reactions were quenched and analyzed by Western blotting for GP1 and GP2. The variable staining intensities of the 130-kDa versus the 19-kDa forms of GP1 could be due to the polyclonal GP1 antibody reacting more strongly with the 19-kDa form or to differing efficiencies of transfer between the high- and low-molecular-mass forms.

Effects of inhibitors and siRNA on infection by protease-treated VSV-GP. VSV-GP was treated with no enzyme, 20 μg/ml CatB plus CatL (CatB+L), or 0.5 mg/ml thermolysin (Therm) for 20 min as described in the text. (A) Vero cells were treated with vehicle alone (V) or CatB (B) and CatL (L) inhibitors as described in the legend to Fig. 1A (1 μM CatB inhibitor and 4 μM CatL inhibitor) and then infected with mock-treated or protease-treated VSV-GP at an MOI of 0.02 to 0.04 (note that this is a 10-fold lower MOI than that used for experiments in Fig. 1 to enable quantification of enhanced infection). Twenty-four hours postinfection, the percentage of GFP-positive cells was determined by flow cytometry. Data shown are the averages of duplicate infections from one representative experiment. Error bars indicate the standard error. (B and C) Vero cells were treated with CatB (B) and CatL (L) inhibitors (B) or siRNA (C) as described in the legend to Fig. 1 and infected with protease-treated VSV-GP as described for panel A. Results shown are the averages of normalized data from three to five experiments; panel B includes the data presented in panel A. Error bars indicate the standard errors, and asterisks indicate a statistically significant deviation from the mean of respective mock-treated virus infections at a confidence level greater than 98% (*) or 94% (**). We note that stronger inhibition by the CatL chemical inhibitor is seen in panel B than that seen in Fig. 1B. This may be due to the lower MOI used in this experiment. (D) Vero cells were left untreated (U) or were pretreated for 2 h at 37°C with 200 nM bafilomycin (bf) or 50 μM E64d (E) and then infected with protease-treated VSV-GP as described above or with VSV-G at an MOI of 0.5 to 1. Twenty-four hours postinfection, the percentage of GFP-positive cells was determined by fluorescence microscopy. Results shown are the averages of normalized data from two to three experiments, and error bars indicate standard errors.

Models of GP-mediated fusion. (A) In our model, initial cleavage of GP1 (by CatB, CatL, or other proteases) removes the mucin-like domain generating a 50-kDa species (not shown). This 50-kDa species is then primed by CatB plus CatL (or another protease) cleavage of GP1 to produce 20-kDa (royal blue) and then 19-kDa (purple) forms of the glycoprotein. This primed form of GP is then acted upon by an additional cellular factor, such as a lysosomal thiol reductase, which reduces a critical disulfide bond, possibly the disulfide bond between GP1 and GP2. This reduction relieves the GP1 clamp, thus allowing conformational changes in GP2 that expose and reposition the fusion peptide (red arrowhead) and trigger fusion. (B) In the model proposed by Chandran et al. (2), initial cleavages by CatB and/or CatL generate an 18-kDa GP1 intermediate (royal blue, which we hypothesize is equivalent to our 20-kDa intermediate with one N-linked glycosylation site removed). Further cleavage by CatB completely digests GP1, thereby removing the clamp and allowing fusion. N-linked glycosylation sites in GP1 are depicted with short branched black lines.
References
-
- Ebert, D. H., J. Deussing, C. Peters, and T. S. Dermody. 2002. Cathepsin L and cathepsin B mediate reovirus disassembly in murine fibroblast cells. J. Biol. Chem. 277:24609-24617. - PubMed
-
- Golden, J., and L. Schiff. 2005. Neutrophil elastase, an acid-independent serine protease, facilitates reovirus uncoating and infection in U937 promonocyte cells. Virol. J. 2:48. http://www.virologyj.com/content/2/1/48. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
