

Bip is a molecular link between the phase I and phase II estrogenic responses in uterus
- PMID: 16574737
- PMCID: PMC4269476
- DOI: 10.1210/me.2006-0046
Bip is a molecular link between the phase I and phase II estrogenic responses in uterus
Abstract
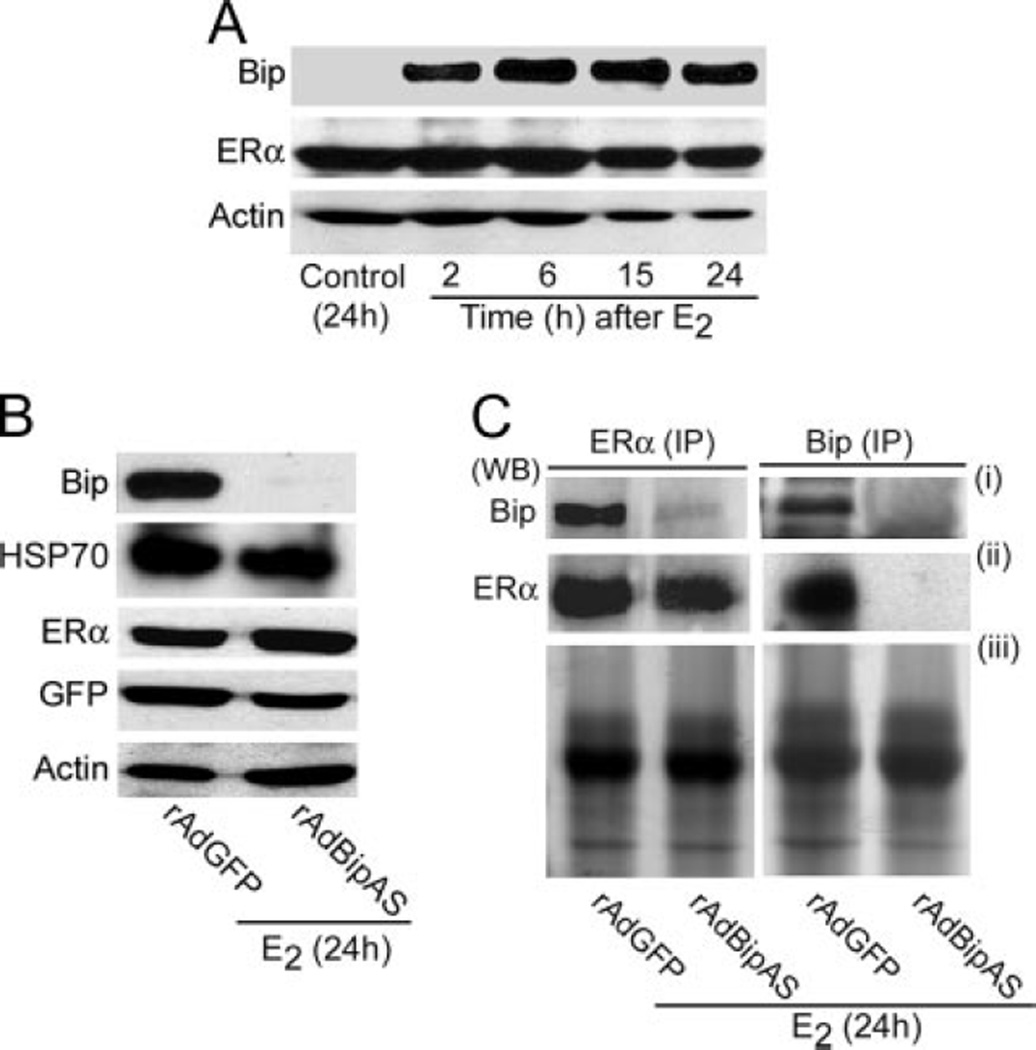
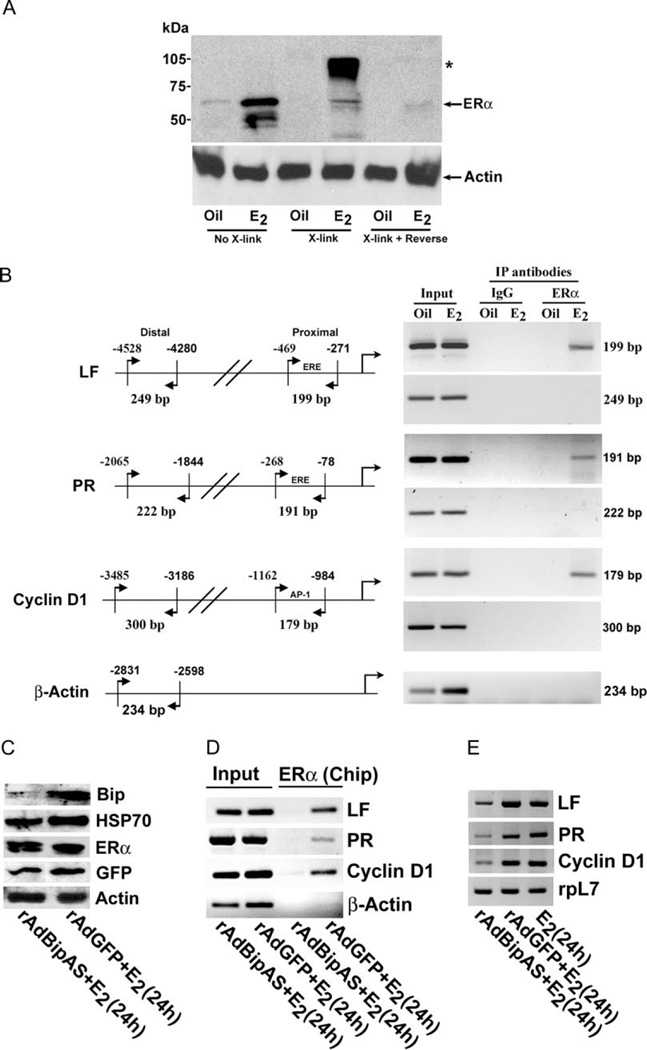
Uterine estrogenic actions are biphasic, early (phase I) and late (phase II) responses. However, the molecular linkage between these phases is not known. Although certain phase I responses are considered estrogen receptor (ER)alpha and ERbeta independent, the phase II responses are ERalpha dependent. We previously observed that among several genes Bip is induced by estrogen in the mouse uterus in an ER-independent manner as a phase I response. Bip is a member of the chaperone family and plays roles in protein processing and confers cellular protection. However, its role in estrogen-dependent uterine biology is unknown. We show here a new function of Bip in regulating estrogen signaling in the uterus. Bip, induced during the phase I responses, molecularly interacts with ERalpha required for estrogen-mediated phase II growth responses. Utilizing in vivo and in vitro model systems, we found that adenovirus-driven suppression of Bip antagonizes ERalpha-mediated uterine gene transcription. Importantly, down-regulation of Bip compromises estrogen-dependent phase II growth responses with sustained phase I responses. In conclusion, Bip is critical for coordinating estrogen-elicited biphasic responses and serves as a molecular link between ERalpha-independent and -dependent estrogenic responses in the uterus.
Figures




Similar articles
-
Increased level of cellular Bip critically determines estrogenic potency for a xenoestrogen kepone in the mouse uterus.Endocrinology. 2007 Oct;148(10):4774-85. doi: 10.1210/en.2007-0537. Epub 2007 Jul 19. Endocrinology. 2007. PMID: 17640991 Free PMC article.
-
Estrogen targets genes involved in protein processing, calcium homeostasis, and Wnt signaling in the mouse uterus independent of estrogen receptor-alpha and -beta.J Biol Chem. 2000 Sep 15;275(37):28834-42. doi: 10.1074/jbc.M003827200. J Biol Chem. 2000. PMID: 10893236
-
Estren behaves as a weak estrogen rather than a nongenomic selective activator in the mouse uterus.Endocrinology. 2006 May;147(5):2203-14. doi: 10.1210/en.2005-1292. Epub 2006 Feb 9. Endocrinology. 2006. PMID: 16469803
-
Differential spatiotemporal regulation of lactoferrin and progesterone receptor genes in the mouse uterus by primary estrogen, catechol estrogen, and xenoestrogen.Endocrinology. 1998 Jun;139(6):2905-15. doi: 10.1210/endo.139.6.6051. Endocrinology. 1998. PMID: 9607801 Free PMC article.
-
Estrogen signaling: a subtle balance between ER alpha and ER beta.Mol Interv. 2003 Aug;3(5):281-92. doi: 10.1124/mi.3.5.281. Mol Interv. 2003. PMID: 14993442 Review.
Cited by
-
GRP78 expression and regulation in the mouse uterus during embryo implantation.J Mol Histol. 2014 Jun;45(3):259-68. doi: 10.1007/s10735-013-9552-1. Epub 2013 Nov 16. J Mol Histol. 2014. PMID: 24242779
-
Heat Shock Proteins and Their Role in Pregnancy: Redefining the Function of "Old Rum in a New Bottle".Front Cell Dev Biol. 2021 Apr 29;9:648463. doi: 10.3389/fcell.2021.648463. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33996811 Free PMC article. Review.
-
Cooperative control via lymphoid enhancer factor 1/T cell factor 3 and estrogen receptor-alpha for uterine gene regulation by estrogen.Mol Endocrinol. 2008 May;22(5):1125-40. doi: 10.1210/me.2007-0445. Epub 2008 Jan 17. Mol Endocrinol. 2008. PMID: 18202148 Free PMC article.
-
Mouse primary uterine cell coculture system revisited: ovarian hormones mimic the aspects of in vivo uterine cell proliferation.Endocrinology. 2011 Aug;152(8):3246-58. doi: 10.1210/en.2011-0223. Epub 2011 Jun 21. Endocrinology. 2011. PMID: 21693674 Free PMC article.
-
GPR30 activation opposes estrogen-dependent uterine growth via inhibition of stromal ERK1/2 and estrogen receptor alpha (ERα) phosphorylation signals.Endocrinology. 2011 Apr;152(4):1434-47. doi: 10.1210/en.2010-1368. Epub 2011 Feb 8. Endocrinology. 2011. PMID: 21303939 Free PMC article.
References
-
- Couse JF, Korach KS. Estrogen receptor null mice: what have we learned and where will they lead us? Endocr Rev. 1999;20:358–417. - PubMed
-
- Tsai MJ, O’Malley BW. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem. 1994;63:451–486. - PubMed
-
- Dey SK, Lim H, Das SK, Reese J, Paria BC, Daikoku T, Wang H. Molecular cues to implantation. Endocr Rev. 2004;25:341–373. - PubMed
-
- Das SK, Tan J, Raja S, Halder J, Paria BC, Dey SK. Estrogen targets genes involved in protein processing, calcium homeostasis and Wnt signaling in the mouse uterus independent of estrogen receptor-α and -β. J Biol Chem. 2000;275:28834–28842. - PubMed
-
- Hewitt SC, Deroo BJ, Hansen K, Collins J, Grissom S, Afshari CA, Korach KS. Estrogen receptor-dependent genomic responses in the uterus mirror the biphasic physiological response to estrogen. Mol Endocrinol. 2003;17:2070–2083. - PubMed
