

Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells
- PMID: 16585960
- PMCID: PMC1421340
- DOI: 10.1172/JCI22689
Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells
Abstract
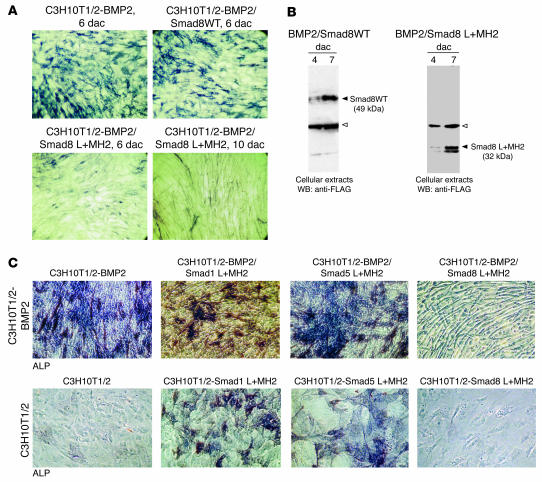
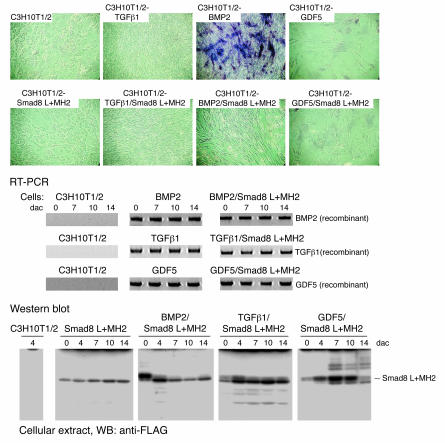
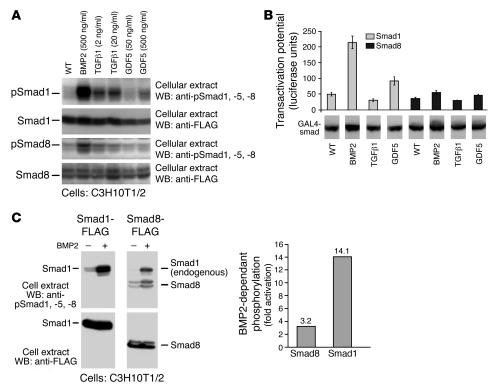
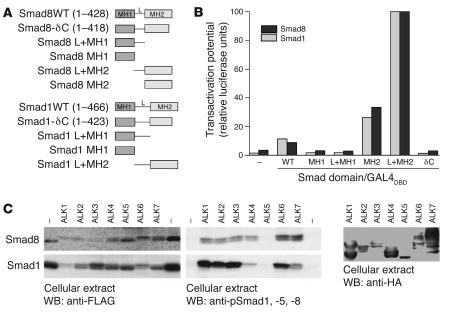
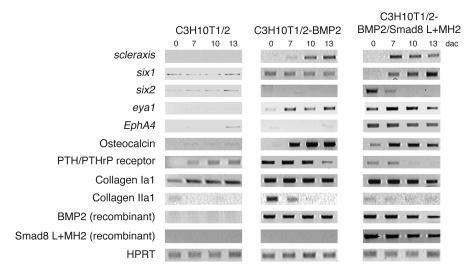
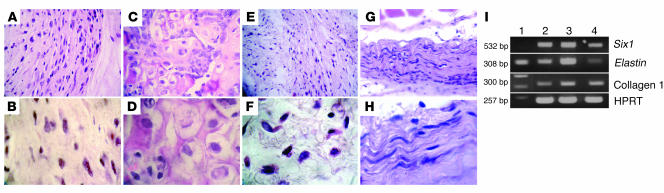
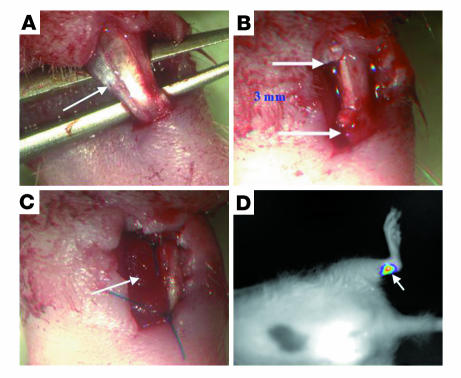
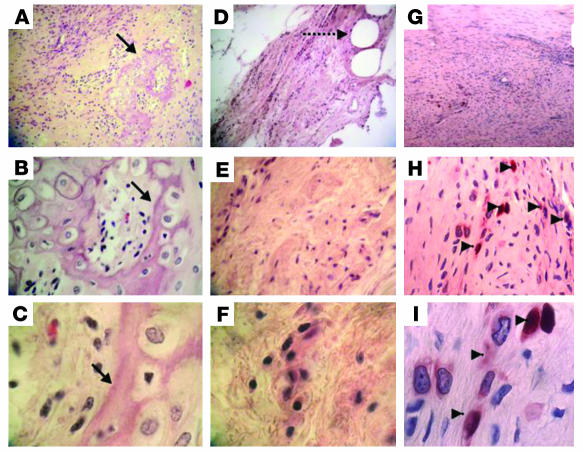
Tissue regeneration requires the recruitment of adult stem cells and their differentiation into mature committed cells. In this study we describe what we believe to be a novel approach for tendon regeneration based on a specific signalling molecule, Smad8, which mediates the differentiation of mesenchymal stem cells (MSCs) into tendon-like cells. A biologically active Smad8 variant was transfected into an MSC line that coexpressed the osteogenic gene bone morphogenetic protein 2 (BMP2). The engineered cells demonstrated the morphological characteristics and gene expression profile of tendon cells both in vitro and in vivo. In addition, following implantation in an Achilles tendon partial defect, the engineered cells were capable of inducing tendon regeneration demonstrated by double quantum filtered MRI. The results indicate what we believe to be a novel mechanism in which Smad8 inhibits the osteogenic pathway in MSCs known to be induced by BMP2 while promoting tendon differentiation. These findings may have considerable importance for the therapeutic replacement of tendons or ligaments and for engineering other tissues in which BMP plays a pivotal developmental role.
Figures
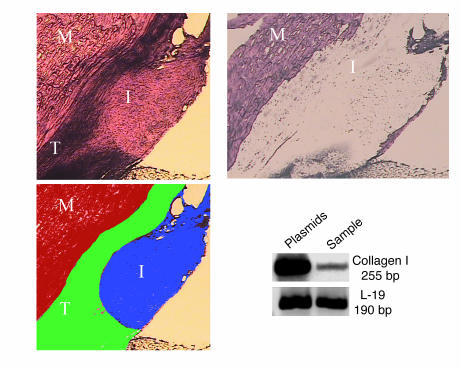
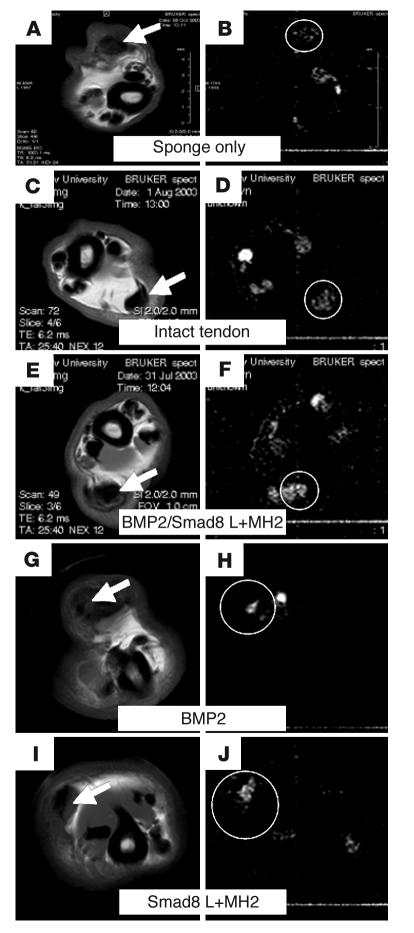
Comment in
-
The alchemy of tendon repair: a primer for the (S)mad scientist.J Clin Invest. 2006 Apr;116(4):863-6. doi: 10.1172/JCI28320. J Clin Invest. 2006. PMID: 16585955 Free PMC article.
References
-
- Pennisi E. Tending tender tendons. Science. 2002;2955:1011. - PubMed
-
- Jackson D.W., Simon T.M., Kurzweil P.R., Rosen M.A. Survival of cells after intra-articular transplantation of fresh allografts of the patellar and anterior cruciate ligaments. DNA-probe analysis in a goat model. J. Bone Joint Surg. Am. 1992;74:112–118. - PubMed
-
- Sabiston P., Frank C., Lam T., Shrive N. Allograft ligament transplantation. A morphological and biochemical evaluation of a medial collateral ligament complex in a rabbit model. Am. J. Sports Med. 1990;18:160–168. - PubMed
-
- Olson E.J., et al. The biochemical and histological effects of artificial ligament wear particles: in vitro and in vivo studies. Am. J. Sports Med. 1988;16:558–570. - PubMed
-
- Woods G.A., Indelicato P.A., Prevot T.J. The Gore-Tex anterior cruciate ligament prosthesis. Two versus three year results. Am. J. Sports Med. 1991;19:48–55. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
