

Ribonuclease P: the evolution of an ancient RNA enzyme
- PMID: 16595295
- PMCID: PMC2803672
- DOI: 10.1080/10409230600602634
Ribonuclease P: the evolution of an ancient RNA enzyme
Abstract
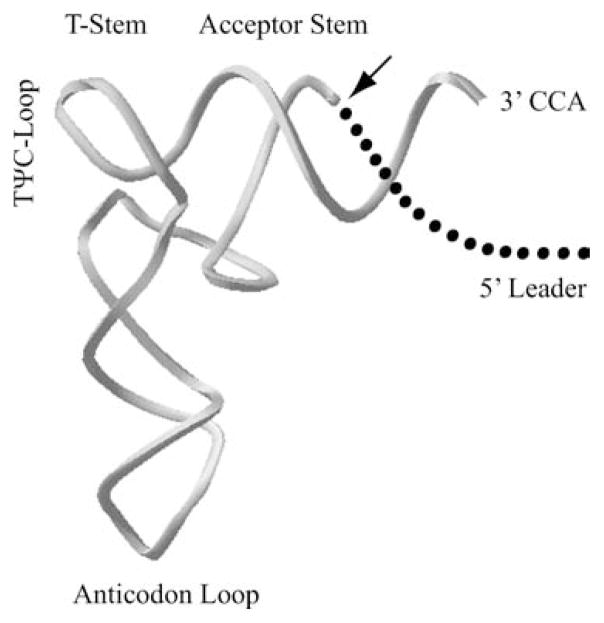
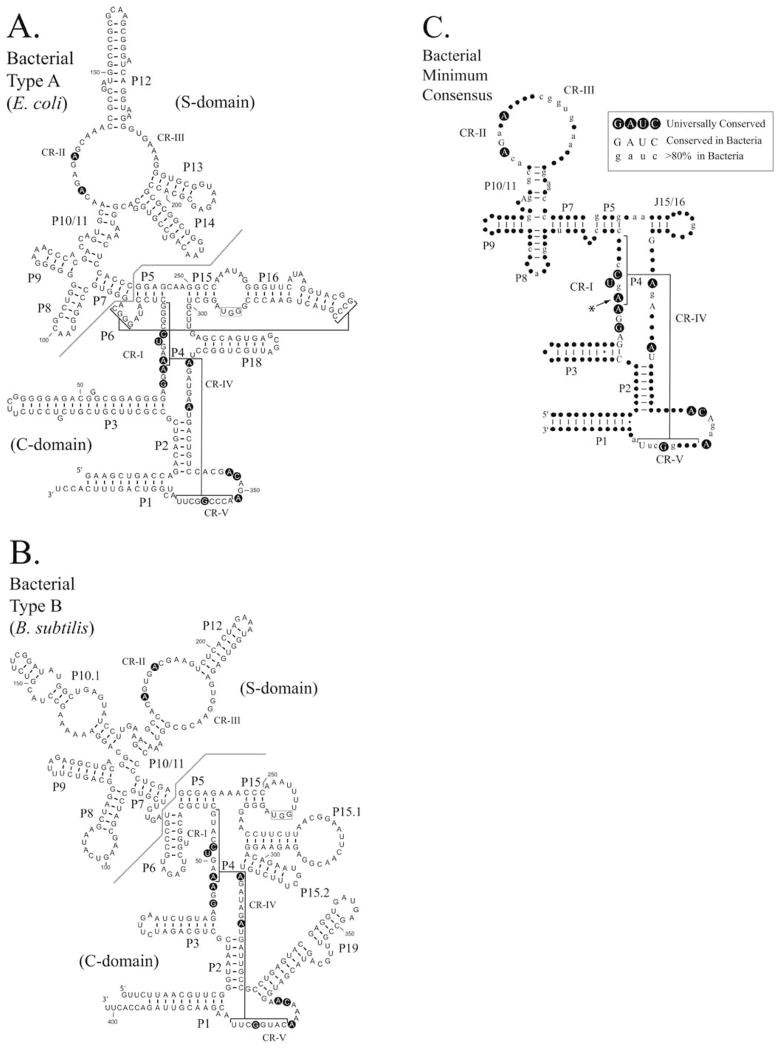
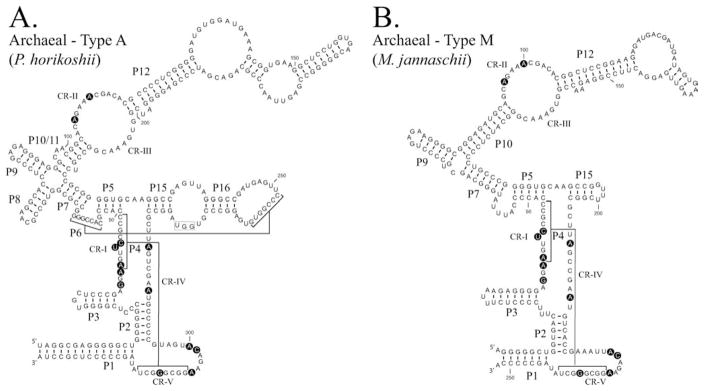
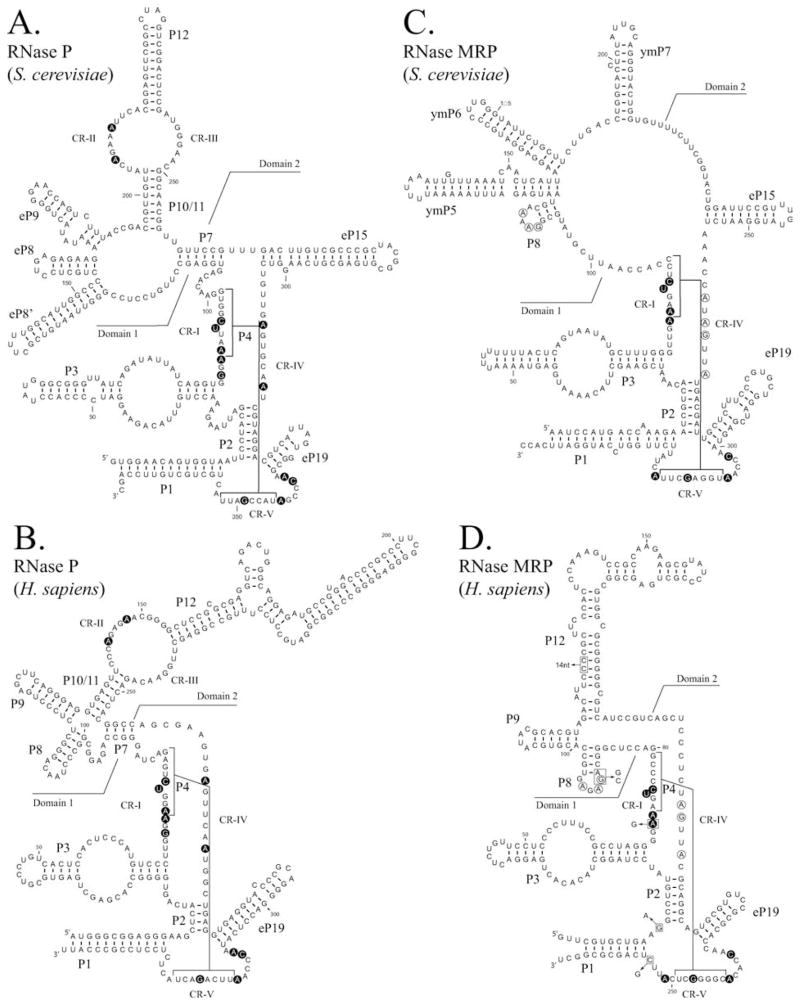
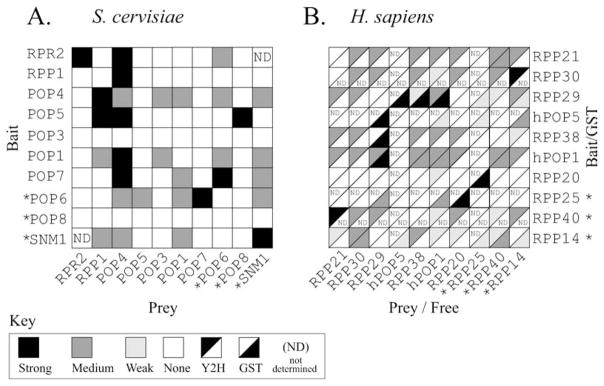
Ribonuclease P (RNase P) is an ancient and essential endonuclease that catalyses the cleavage of the 5' leader sequence from precursor tRNAs (pre-tRNAs). The enzyme is one of only two ribozymes which can be found in all kingdoms of life (Bacteria, Archaea, and Eukarya). Most forms of RNase P are ribonucleoproteins; the bacterial enzyme possesses a single catalytic RNA and one small protein. However, in archaea and eukarya the enzyme has evolved an increasingly more complex protein composition, whilst retaining a structurally related RNA subunit. The reasons for this additional complexity are not currently understood. Furthermore, the eukaryotic RNase P has evolved into several different enzymes including a nuclear activity, organellar activities, and the evolution of a distinct but closely related enzyme, RNase MRP, which has different substrate specificities, primarily involved in ribosomal RNA biogenesis. Here we examine the relationship between the bacterial and archaeal RNase P with the eukaryotic enzyme, and summarize recent progress in characterizing the archaeal enzyme. We review current information regarding the nuclear RNase P and RNase MRP enzymes in the eukaryotes, focusing on the relationship between these enzymes by examining their composition, structure and functions.
Figures
References
-
- Alifano P, Rivellini F, Piscitelli C, Arraiano CM, Bruni CB, Carlomagno MS. Ribonuclease E provides substrates for ribonuclease P-dependent processing of a polycistronic mRNA. Genes Dev. 1994;8(24):3021. - PubMed
-
- Altman S, Wesolowski D, Puranam RS. Nucleotide sequences of the RNA subunit of RNase P from several mammals. Genomics. 1993;18(2):418. - PubMed
-
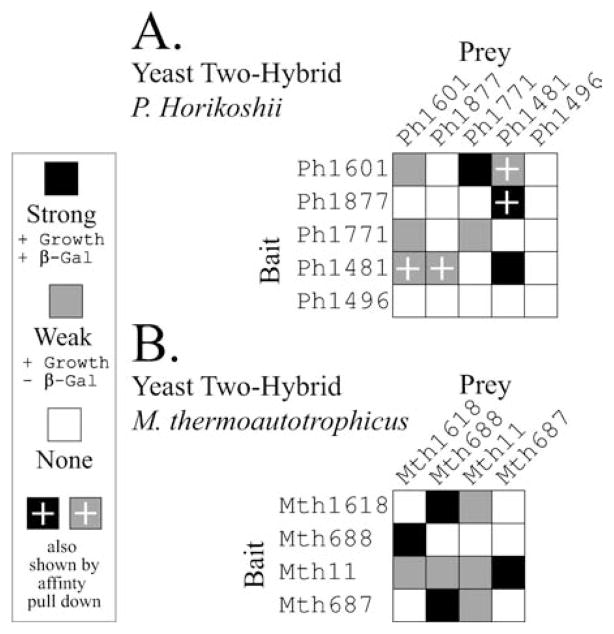
- Andrews AJ, Hall TA, Brown JW. Characterization of RNase P holoenzymes from Methanococcus jannaschii and Methanothermobacter thermoautotrophicus. Biol Chem. 2001;382(8):1171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases