

Modular organization of FDH: Exploring the basis of hydrolase catalysis
- PMID: 16597835
- PMCID: PMC2242502
- DOI: 10.1110/ps.052062806
Modular organization of FDH: Exploring the basis of hydrolase catalysis
Abstract
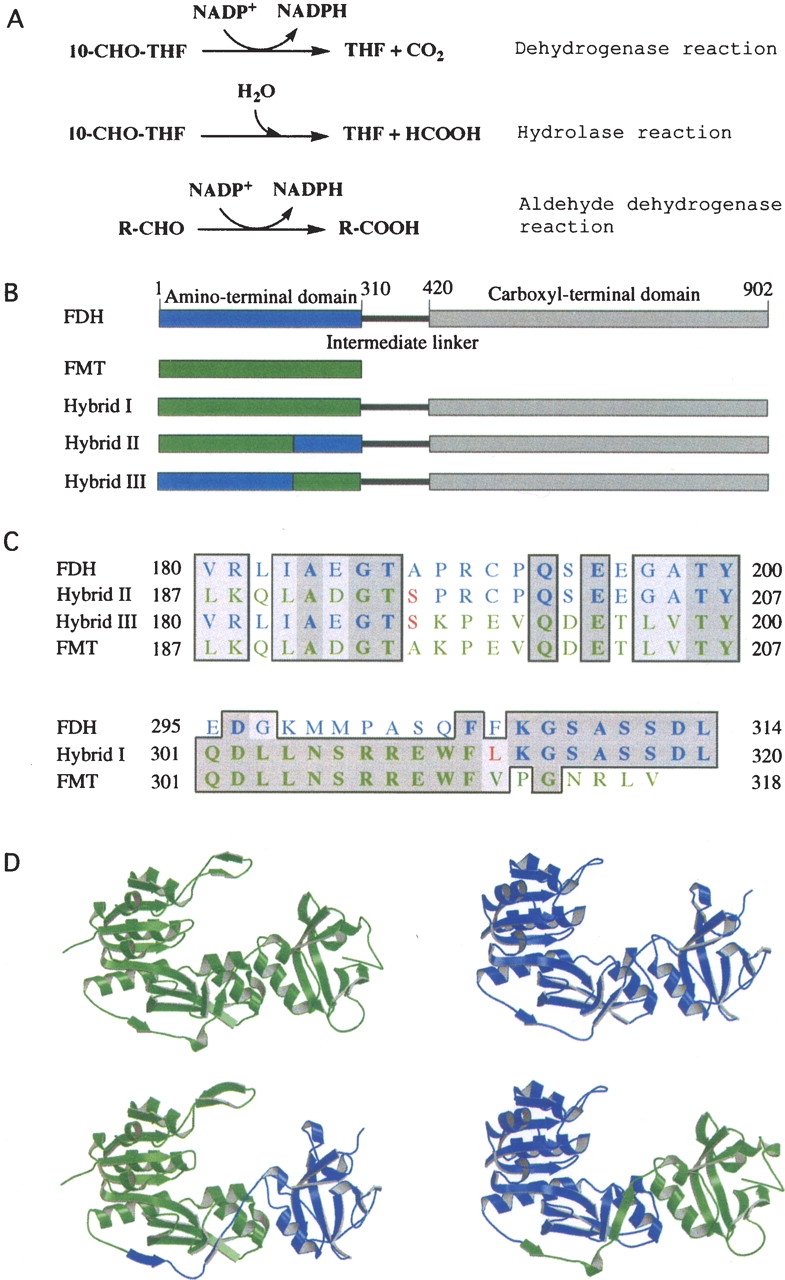
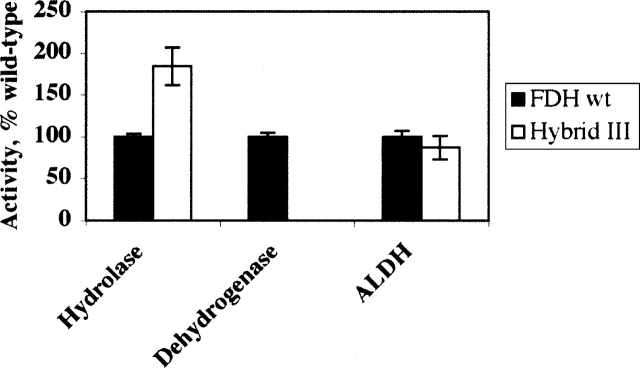
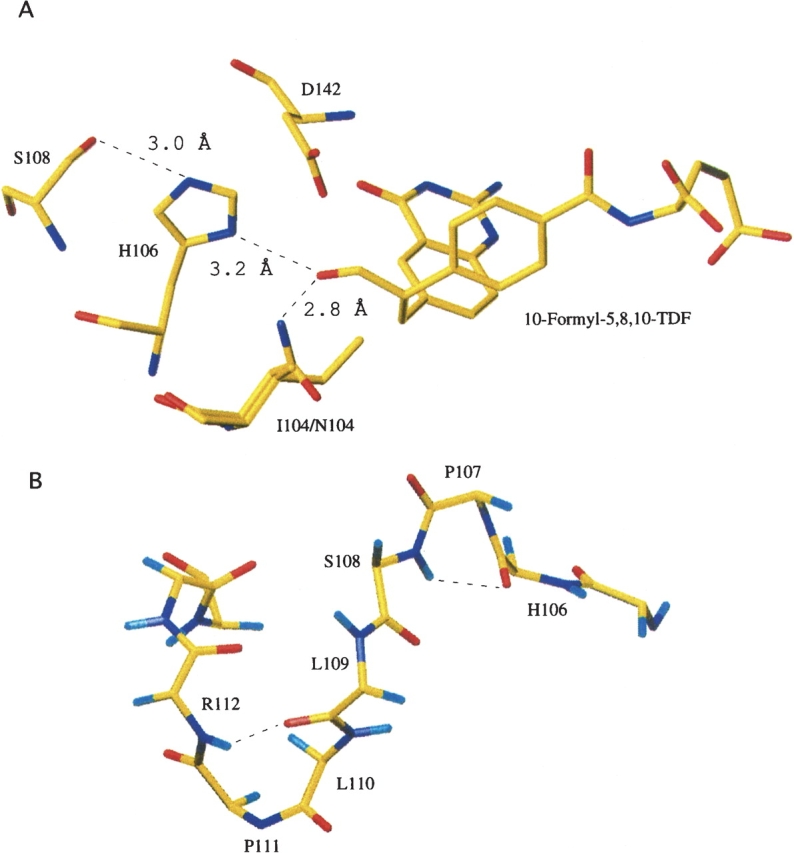
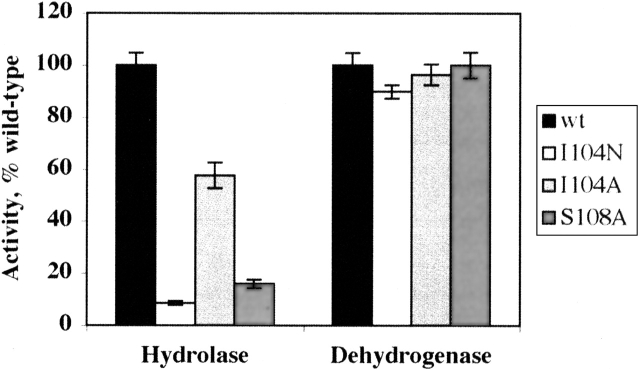
An abundant enzyme of liver cytosol, 10-formyltetrahydrofolate dehydrogenase (FDH), is an interesting example of a multidomain protein. It consists of two functionally unrelated domains, an aldehyde dehydrogenase-homologous domain and a folate-binding hydrolase domain, which are connected by an approximately 100-residue linker. The amino-terminal hydrolase domain of FDH (Nt-FDH) is a homolog of formyl transferase enzymes that utilize 10-formyl-THF as a formyl donor. Interestingly, the concerted action of all three domains of FDH produces a new catalytic activity, NADP+-dependent oxidation of 10-formyltetrahydrofolate (10-formyl-THF) to THF and CO2. The present studies had two objectives: First, to explore the modular organization of FDH through the production of hybrid enzymes by domain replacement with methionyl-tRNA formyltransferase (FMT), an enzyme homologous to the hydrolase domain of FDH. The second was to explore the molecular basis for the distinct catalytic mechanisms of Nt-FDH and related 10-formyl-THF utilizing enzymes. Our studies revealed that FMT cannot substitute for the hydrolase domain of FDH in order to catalyze the dehydrogenase reaction. It is apparently due to inability of FMT to catalyze the hydrolysis of 10-formyl-THF in the absence of the cosubstrate of the transferase reaction despite the high similarity of the catalytic centers of the two enzymes. Our results further imply that Ile in place of Asn in the FDH hydrolase catalytic center is an important determinant for hydrolase catalysis as opposed to transferase catalysis.
Figures
Similar articles
-
Structures of the hydrolase domain of zebrafish 10-formyltetrahydrofolate dehydrogenase and its complexes reveal a complete set of key residues for hydrolysis and product inhibition.Acta Crystallogr D Biol Crystallogr. 2015 Apr;71(Pt 4):1006-21. doi: 10.1107/S1399004715002928. Epub 2015 Mar 27. Acta Crystallogr D Biol Crystallogr. 2015. PMID: 25849409 Free PMC article.
-
On the role of conserved histidine 106 in 10-formyltetrahydrofolate dehydrogenase catalysis: connection between hydrolase and dehydrogenase mechanisms.J Biol Chem. 2001 Jun 29;276(26):24030-7. doi: 10.1074/jbc.M009257200. Epub 2001 Apr 24. J Biol Chem. 2001. PMID: 11320079
-
FDH: an aldehyde dehydrogenase fusion enzyme in folate metabolism.Chem Biol Interact. 2009 Mar 16;178(1-3):84-93. doi: 10.1016/j.cbi.2008.09.007. Epub 2008 Sep 19. Chem Biol Interact. 2009. PMID: 18848533 Free PMC article. Review.
-
The crystal structure of the hydrolase domain of 10-formyltetrahydrofolate dehydrogenase: mechanism of hydrolysis and its interplay with the dehydrogenase domain.J Biol Chem. 2004 Apr 2;279(14):14355-64. doi: 10.1074/jbc.M313934200. Epub 2004 Jan 16. J Biol Chem. 2004. PMID: 14729668
-
Enzyme promiscuity in the hormone-sensitive lipase family of proteins.Protein Pept Lett. 2012 Feb;19(2):144-54. doi: 10.2174/092986612799080400. Protein Pept Lett. 2012. PMID: 21933124 Review.
Cited by
-
Acyl carrier protein-specific 4'-phosphopantetheinyl transferase activates 10-formyltetrahydrofolate dehydrogenase.J Biol Chem. 2010 Jan 15;285(3):1627-33. doi: 10.1074/jbc.M109.080556. Epub 2009 Nov 20. J Biol Chem. 2010. PMID: 19933275 Free PMC article.
-
Loss of ALDH1L1 folate enzyme confers a selective metabolic advantage for tumor progression.Chem Biol Interact. 2019 Apr 1;302:149-155. doi: 10.1016/j.cbi.2019.02.013. Epub 2019 Feb 20. Chem Biol Interact. 2019. PMID: 30794800 Free PMC article. Review.
-
Modeling of interactions between functional domains of ALDH1L1.Chem Biol Interact. 2017 Oct 1;276:23-30. doi: 10.1016/j.cbi.2017.04.011. Epub 2017 Apr 14. Chem Biol Interact. 2017. PMID: 28414156 Free PMC article.
-
Structures of the hydrolase domain of zebrafish 10-formyltetrahydrofolate dehydrogenase and its complexes reveal a complete set of key residues for hydrolysis and product inhibition.Acta Crystallogr D Biol Crystallogr. 2015 Apr;71(Pt 4):1006-21. doi: 10.1107/S1399004715002928. Epub 2015 Mar 27. Acta Crystallogr D Biol Crystallogr. 2015. PMID: 25849409 Free PMC article.
-
Further delineation of the phenotypic and metabolomic profile of ALDH1L2-related neurodevelopmental disorder.Clin Genet. 2024 May;105(5):488-498. doi: 10.1111/cge.14479. Epub 2024 Jan 9. Clin Genet. 2024. PMID: 38193334 Free PMC article.
References
-
- Bhatt A.N., Khan M.Y., Bhakuni V. 2004. The C-terminal domain of dimeric serine hydroxymethyltransferase plays a key role in stabilization of the quaternary structure and cooperative unfolding of protein: Domain swapping studies with enzymes having high sequence identity Protein Sci. 13: 2184–2195. - PMC - PubMed
-
- Chumanevich A.A., Krupenko S.A., Davies C. 2004. The crystal structure of the hydrolase domain of 10-formyltetrahydrofolate dehydrogenase: Mechanism of hydrolysis and its interplay with the dehydrogenase domain J. Biol. Chem. 279: 14355–14364. - PubMed
-
- Cook R.J. and Wagner C. 1995. Enzymatic activities of rat liver cytosol 10-formyltetrahydrofolate dehydrogenase Arch. Biochem. Biophys. 321: 336–344. - PubMed
-
- Cook R.J., Lloyd R.S., Wagner C. 1991. Isolation and characterization of cDNA clones for rat liver 10-formyltetrahydrofolate dehydrogenase J. Biol. Chem. 266: 4965–4973. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
