

The role of Rat1 in coupling mRNA 3'-end processing to transcription termination: implications for a unified allosteric-torpedo model
- PMID: 16598041
- PMCID: PMC1472303
- DOI: 10.1101/gad.1409106
The role of Rat1 in coupling mRNA 3'-end processing to transcription termination: implications for a unified allosteric-torpedo model
Abstract
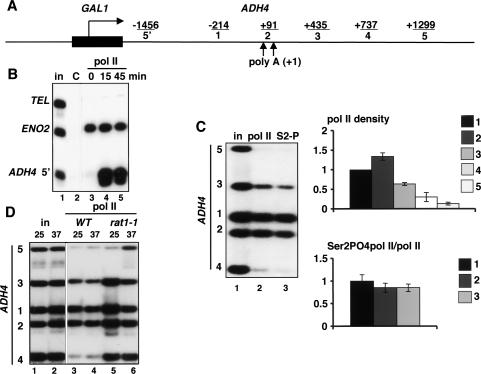
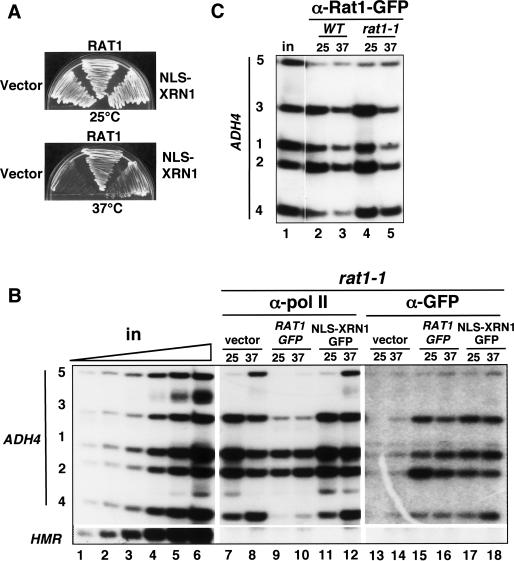
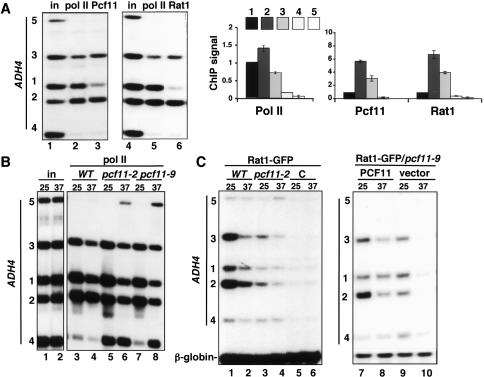
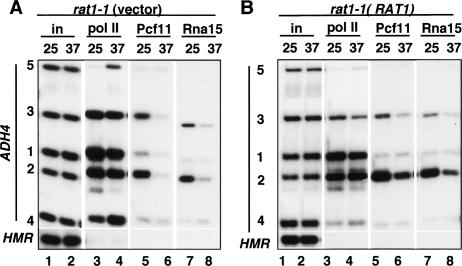
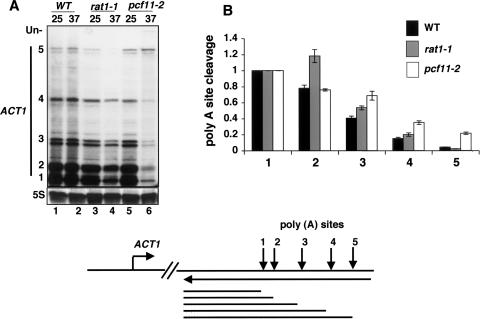
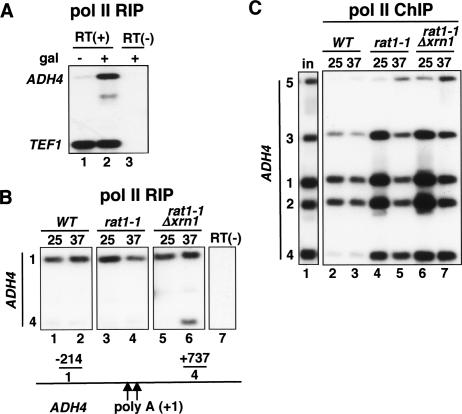
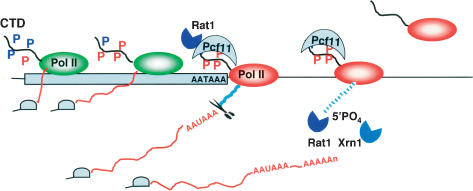
The torpedo model of transcription termination by RNA polymerase II proposes that a 5'-3' RNA exonuclease enters at the poly(A) cleavage site, degrades the nascent RNA, and eventually displaces polymerase from the DNA. Cotranscriptional degradation of nascent RNA has not been directly demonstrated, however. Here we report that two exonucleases, Rat1 and Xrn1, both contribute to cotranscriptional degradation of nascent RNA, but this degradation is not sufficient to cause polymerase release. Unexpectedly, Rat1 functions in both 3'-end processing and termination by enhancing recruitment of 3'-end processing factors, including Pcf11 and Rna15. In addition, the cleavage factor Pcf11 reciprocally aids in recruitment of Rat1 to the elongation complex. Our results suggest a unified allosteric/torpedo model in which Rat1 is not a dedicated termination factor, but is an integrated component of the cleavage/polyadenylation apparatus.
Figures
Comment in
-
Terminating the transcript: breaking up is hard to do.Genes Dev. 2006 May 1;20(9):1050-6. doi: 10.1101/gad.1431606. Genes Dev. 2006. PMID: 16651651 Review. No abstract available.
References
-
- Ahn S.H., Kim M., Buratowski S. Phosphorylation of serine 2 within the RNA polymerase II C-terminal domain couples transcription and 3′ end processing. Mol. Cell. 2004;13:67–76. - PubMed
-
- Amberg D.C., Goldstein A.L., Cole C.N. Isolation and characterization of RAT1: An essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes & Dev. 1992;6:1173–1189. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases