

The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis
- PMID: 16600915
- PMCID: PMC1472290
- DOI: 10.1101/gad.373506
The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis
Abstract
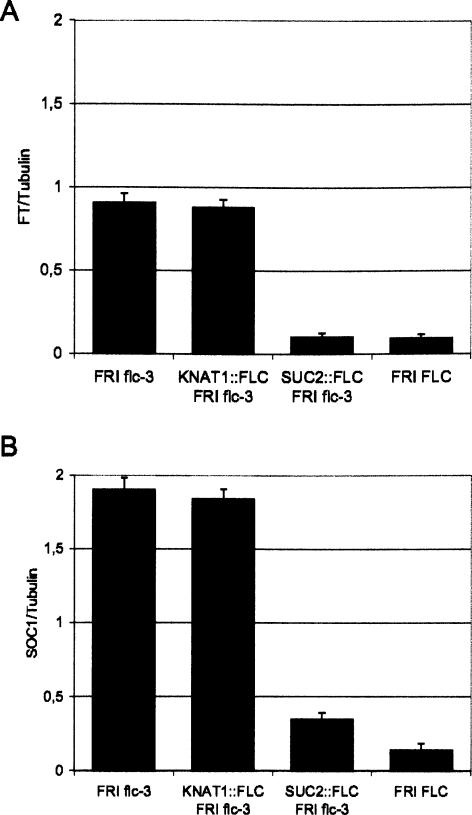
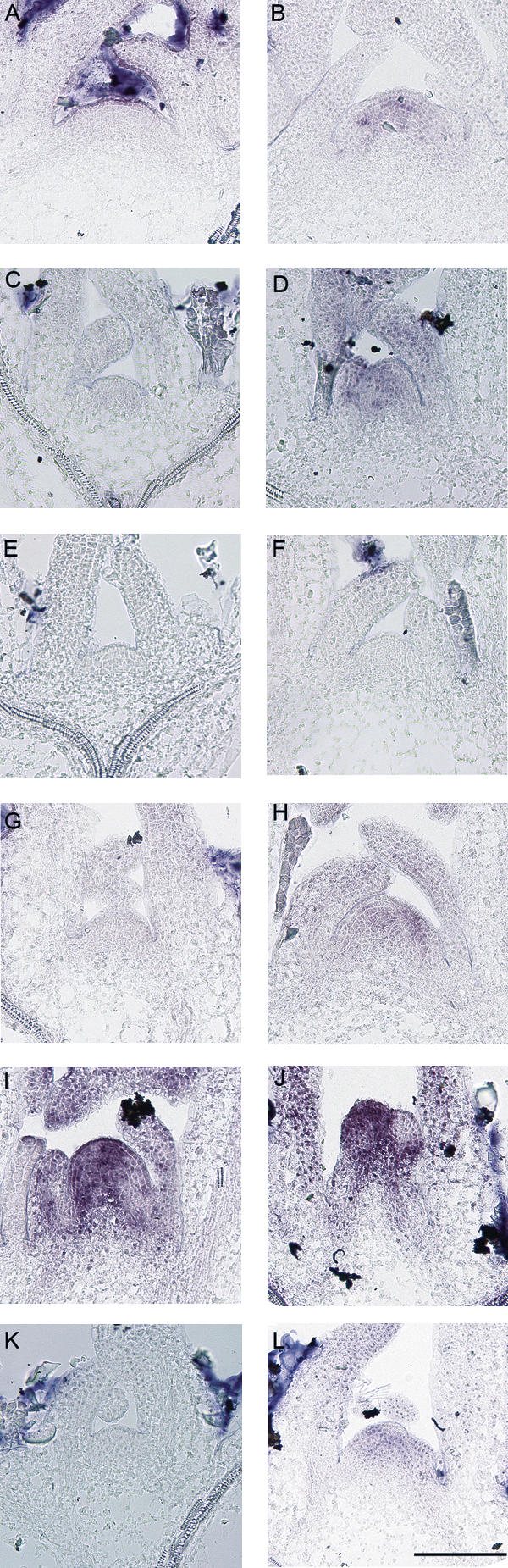
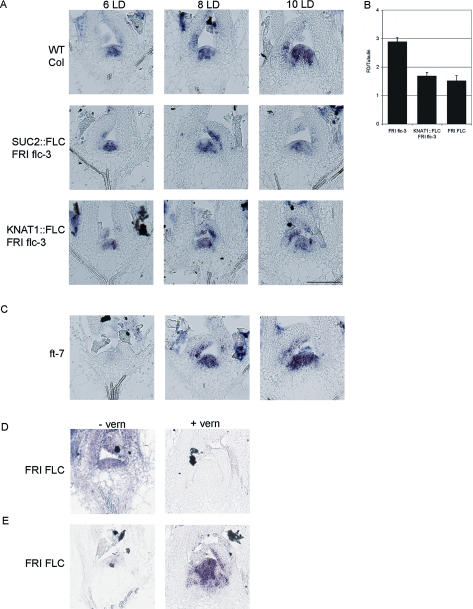
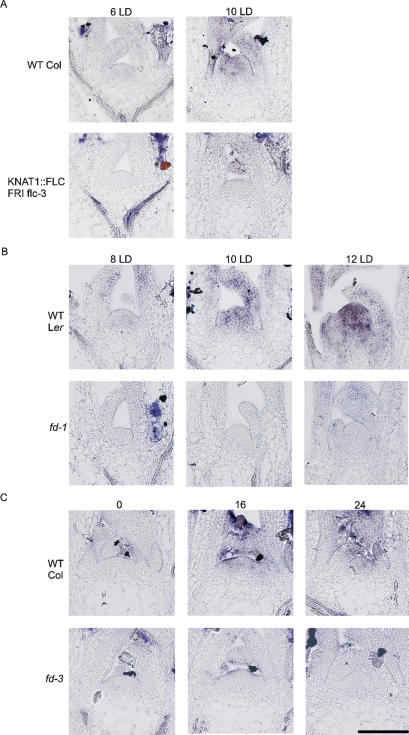
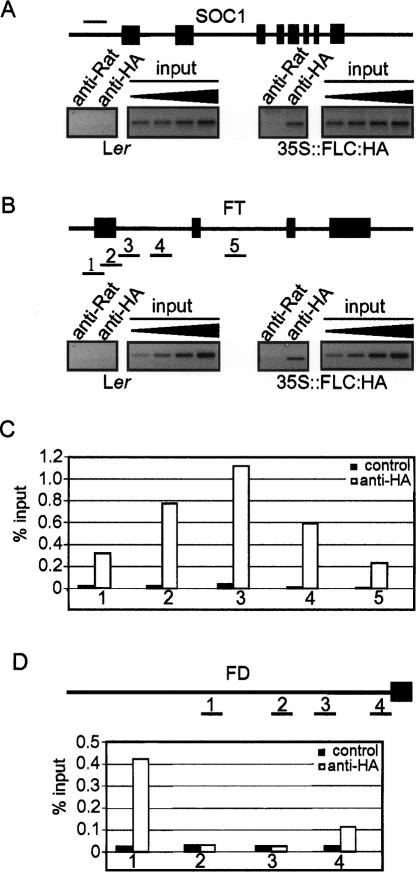
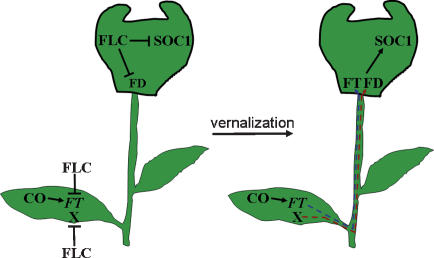
Floral development at the Arabidopsis shoot apical meristem occurs in response to environmental cues that are perceived in different tissues. Photoperiod is detected in the vascular tissue of the leaf (phloem) and promotes production of a systemic signal that induces flowering at the meristem. Vernalization, the response to winter temperatures, overcomes a block on photoperiodic floral induction. In Arabidopsis, this block is caused by inhibitors of flowering that comprise several related MADS-box transcription factors, the most prominent of which is FLC. We show that FLC delays flowering by repressing production in the leaf of at least two systemic signals, one of which is controlled by the RAF kinase inhibitor-like protein FT. Reducing expression of these signals indirectly represses expression of genes involved in floral induction at the meristem. In addition, FLC expression in the meristem impairs response to the FT signal by directly repressing expression of the SOC1 MADS-box transcription factor and preventing up-regulation of the bZIP transcription factor FD. Repression of genes acting at multiple levels in this hierarchy is required for the extreme delay in flowering caused by FLC. An FLC:HA fusion protein binds directly in vivo to the promoter regions of FD and SOC1 and to the first intron of FT. Thus vernalization relieves transcriptional repression of key regulatory genes in both the leaf and meristem, allowing production of systemic signals in the leaves and conferring competence on the meristem to respond to these signals.
Figures


References
-
- Abe M., Kobayashi Y., Yamamoto S., Daimon Y., Yamaguchi A., Ikeda Y., Ichinoki H., Notaguchi M., Goto K., Araki T. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science. 2005;309:1052–1056. - PubMed
-
- An H., Roussot C., Suarez-Lopez P., Corbesier L., Vincent C., Pineiro M., Hepworth S., Mouradov A., Justin S., Turnbull C.G.N., et al. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development. 2004;131:3615–3626. - PubMed
-
- Bastow R., Mylne J.S., Lister C., Lippman Z., Martienssen R.A., Dean C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature. 2004;427:164–167. - PubMed
-
- Blazquez M.A., Weigel D. Integration of floral inductive signals in Arabidopsis. . Nature. 2000;404:889–892. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous