

Chaperone-mediated assembly of centromeric chromatin in vitro
- PMID: 16601098
- PMCID: PMC1431717
- DOI: 10.1073/pnas.0601686103
Chaperone-mediated assembly of centromeric chromatin in vitro
Abstract
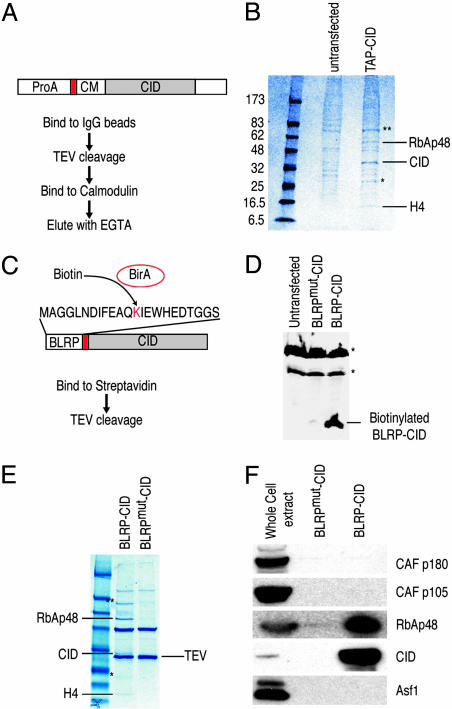
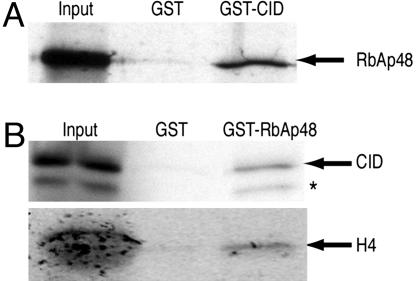
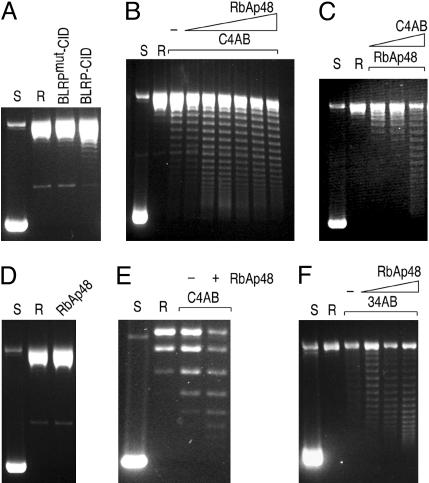
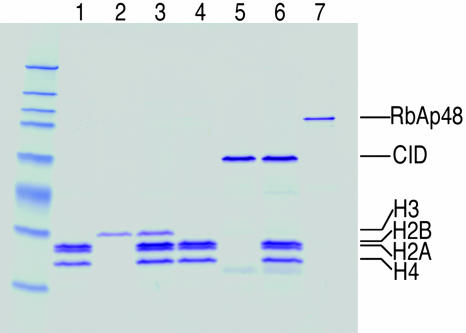
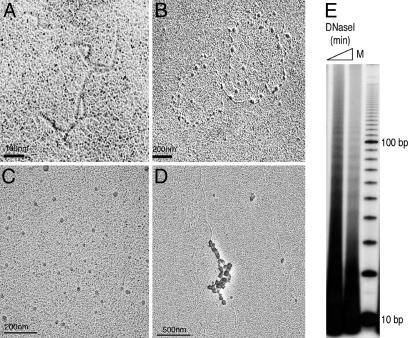
Every eukaryotic chromosome requires a centromere for attachment to spindle microtubules for chromosome segregation. Although centromeric DNA sequences vary greatly among species, centromeres are universally marked by the presence of a centromeric histone variant, centromeric histone 3 (CenH3), which replaces canonical histone H3 in centromeric nucleosomes. Conventional chromatin is maintained in part by histone chaperone complexes, which deposit the S phase-limited (H3) and constitutive (H3.3) forms of histone 3. However, the mechanism that deposits CenH3 specifically at centromeres and faithfully maintains its chromosome location through mitosis and meiosis is unknown. To address this problem, we have biochemically purified a soluble assembly complex that targets tagged CenH3 to centromeres in Drosophila cells. Two different affinity procedures led to purification of the same complex, which consists of CenH3, histone H4, and a single protein chaperone, RbAp48, a highly abundant component of various chromatin assembly, remodeling, and modification complexes. The corresponding CenH3 assembly complex reconstituted in vitro is sufficient for chromatin assembly activity, without requiring additional components. The simple CenH3 assembly complex is in contrast to the multisubunit complexes previously described for H3 and H3.3, suggesting that centromeres are maintained by a passive mechanism that involves exclusion of the complexes that deposit canonical H3s during replication and transcription.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Choo K. H. The Centromere. London: Oxford Univ. Press; 1997.
-
- Malik H. S., Henikoff S. Curr. Opin. Genet. Dev. 2002;12:711–718. - PubMed
-
- Nagaki K., Cheng Z., Ouyang S., Talbert P. B., Kim M., Jones K. M., Henikoff S., Buell C. R., Jiang J. Nat. Genet. 2004;36:138–145. - PubMed
-
- Henikoff S., Ahmad K., Malik H. S. Science. 2001;293:1098–1102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
