

Phosphorylation and activation of PINOID by the phospholipid signaling kinase 3-phosphoinositide-dependent protein kinase 1 (PDK1) in Arabidopsis
- PMID: 16601102
- PMCID: PMC1458890
- DOI: 10.1073/pnas.0510283103
Phosphorylation and activation of PINOID by the phospholipid signaling kinase 3-phosphoinositide-dependent protein kinase 1 (PDK1) in Arabidopsis
Abstract
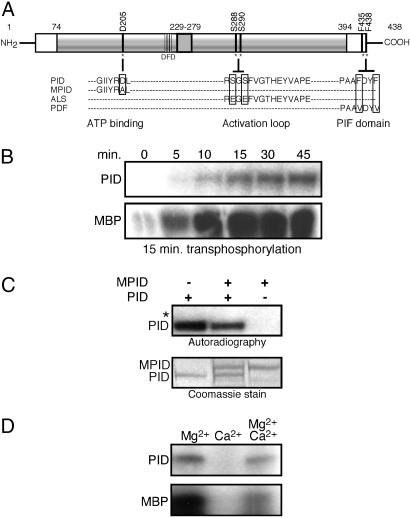
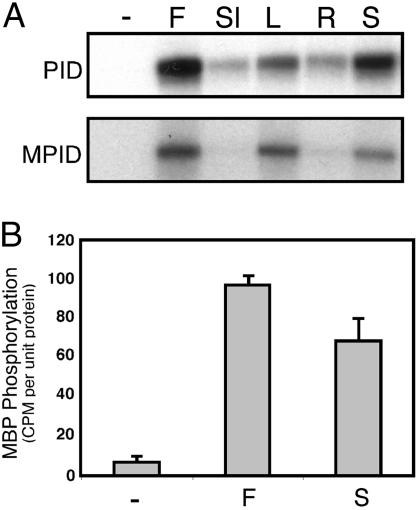
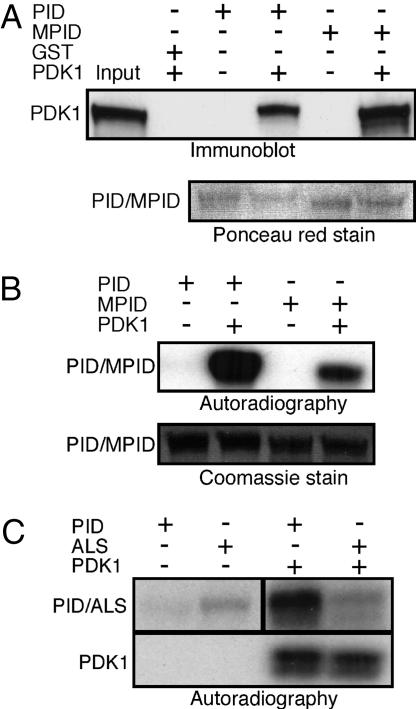
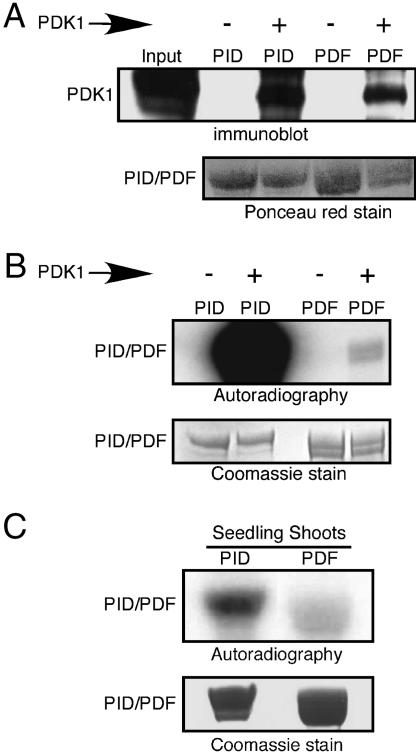
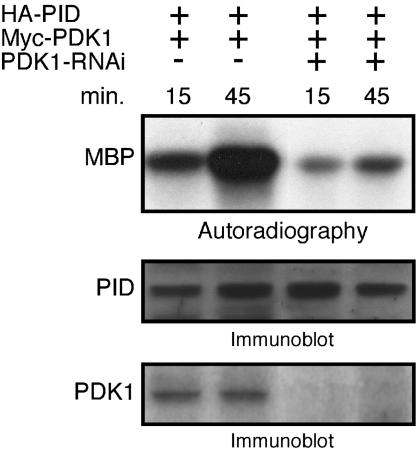
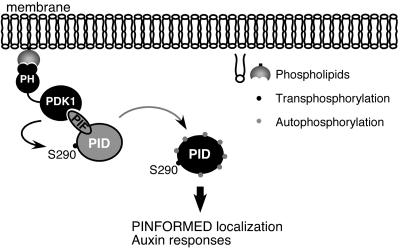
Activity of the serine-threonine protein kinase PINOID (PID) has been implicated in the asymmetrical localization of the membrane-associated PINFORMED (PIN) family of auxin transport facilitators. However, the means by which PID regulates PIN protein distribution is unknown. We have used recombinant PID protein to dissect the regulation of PID activity in vitro. We demonstrate that intramolecular PID autophosphorylation is required for the ability of PID to phosphorylate an exogenous substrate. PID-like mammalian AGC kinases act in a phosphorylation cascade initiated by the phospholipid-associated kinase, 3-phosphoinositide-dependent protein kinase 1 (PDK1), which binds to the C-terminal hydrophobic PDK1-interacting fragment (PIF) domain found in PDK1 substrates. We find that Arabidopsis PDK1 interacts with PID, and that transphosphorylation by PDK1 increases PID autophosphorylation. We show that a PID activation loop serine is required for PDK1-dependent PID phosphorylation. This activation is rapid and requires the PIF domain. Cell extracts from flowers and seedling shoots dramatically increase PID phosphorylation in a tissue-specific manner. A PID protein variant in which the PIF domain was mutated failed to be activated by the seedling shoot extracts. PID immunoprecipitated from Arabidopsis cells in which PDK1 expression was inhibited by RNAi showed a dramatic reduction in transphosphorylation of myelin basic protein substrate. These results indicate that AtPDK1 is a potent enhancer of PID activity and provide evidence that phospholipid signaling may play a role in the signaling processes controlling polar auxin transport.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Friml J., Palme K. Plant Mol. Biol. 2002;49:273–284. - PubMed
-
- Gälweiler L., Guan C., Müller A., Wisman E., Mendgen K., Yephremov A., Palme K. Science. 1998;282:2226–2230. - PubMed
-
- Steinmann T., Geldner N., Grebe M., Mangold S., Jackson C. L., Paris S., Gälweiler L., Palme K., Jürgens G. Science. 1999;286:316–318. - PubMed
-
- Geldner N., Friml J., Stierhof Y. D., Jurgens G., Palme K. Nature. 2001;413:425–428. - PubMed
-
- Geldner N., Anders N., Wolters H., Keicher J., Kornberger W., Muller P., Delbarre A., Ueda T., Nakano A., Jürgens G. Cell. 2003;112:219–230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
