

Nucleobase catalysis in the hairpin ribozyme
- PMID: 16601203
- PMCID: PMC1464849
- DOI: 10.1261/rna.11706
Nucleobase catalysis in the hairpin ribozyme
Abstract
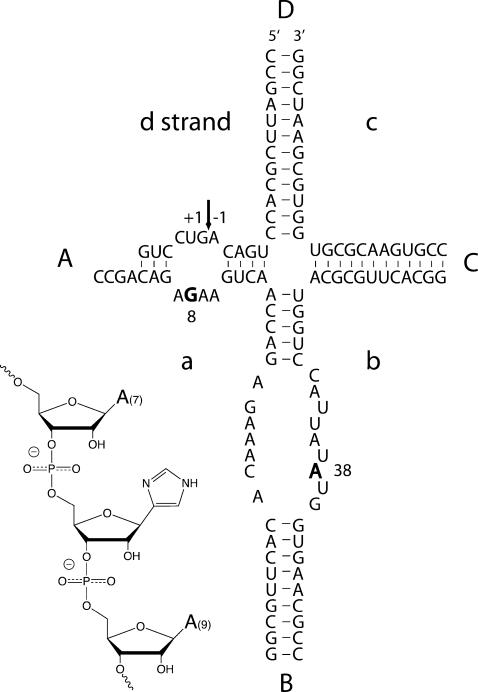
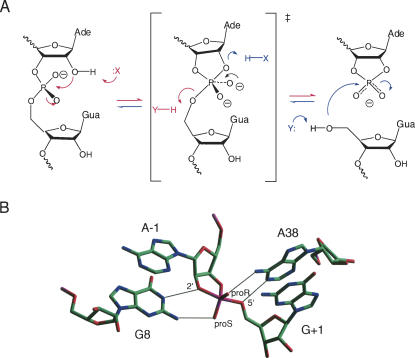
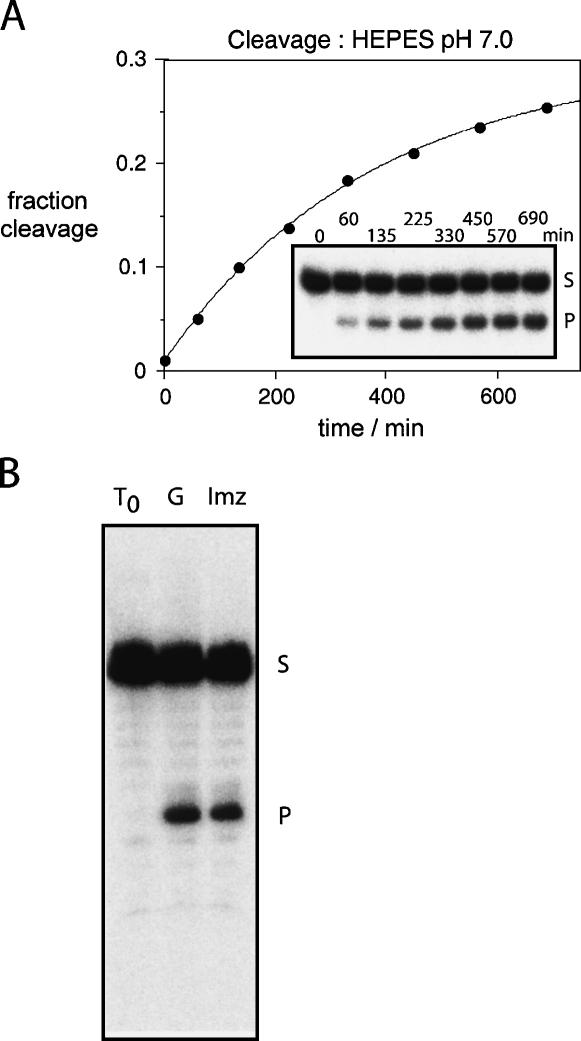
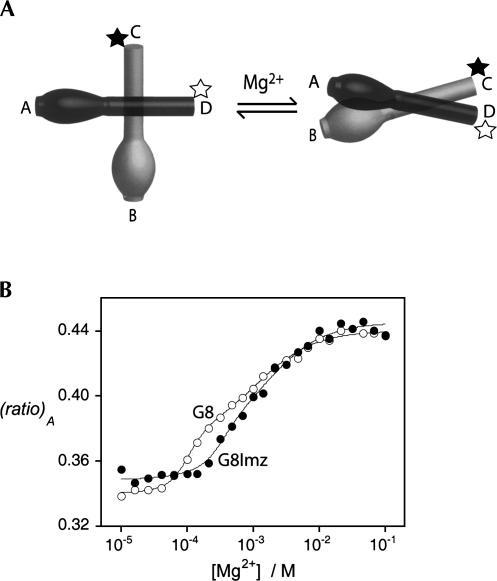
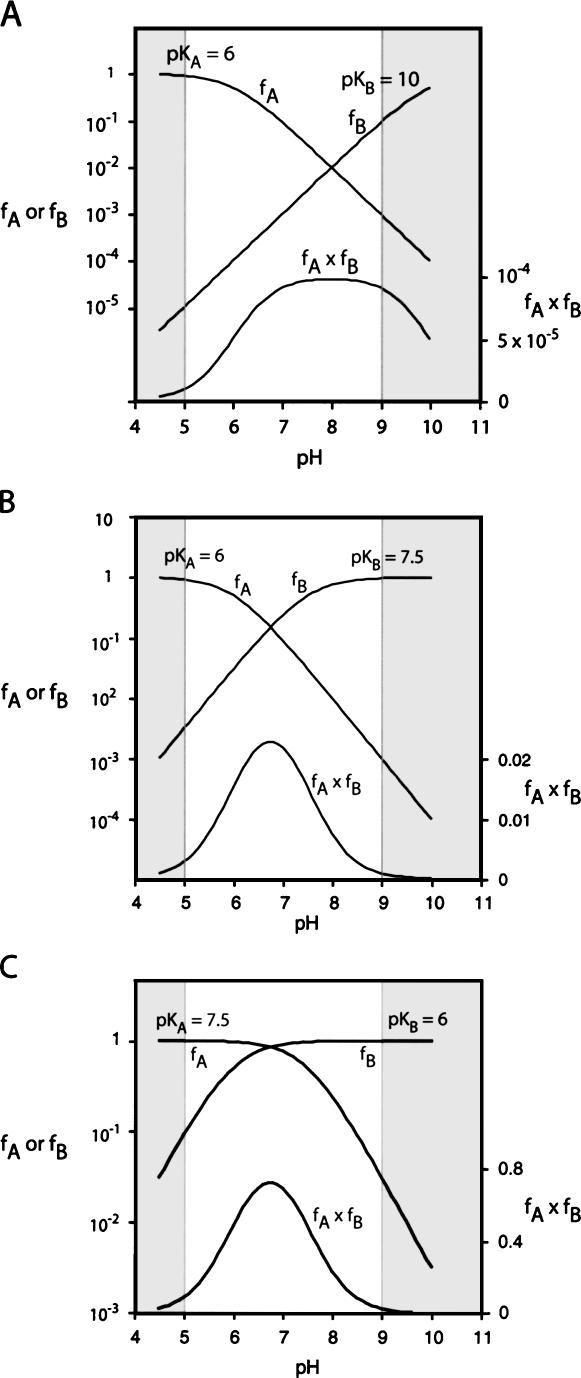
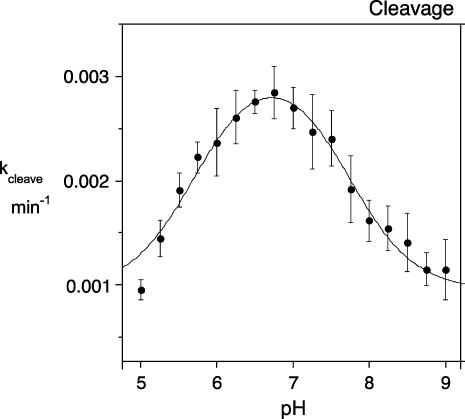
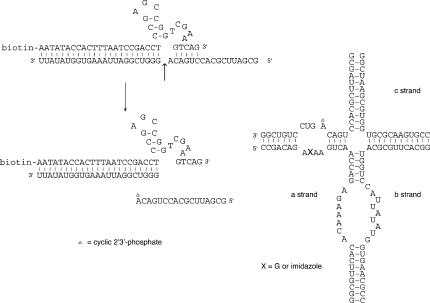
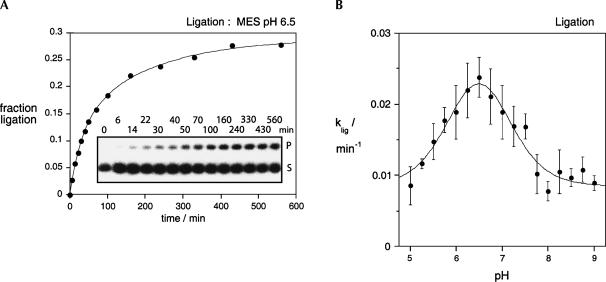
RNA catalysis is important in the processing and translation of RNA molecules, yet the mechanisms of catalysis are still unclear in most cases. We have studied the role of nucleobase catalysis in the hairpin ribozyme, where the scissile phosphate is juxtaposed between guanine and adenine bases. We show that a modified ribozyme in which guanine 8 has been substituted by an imidazole base is active in both cleavage and ligation, with ligation rates 10-fold faster than cleavage. The rates of both reactions exhibit bell-shaped dependence on pH, with pK(a) values of 5.7 +/- 0.1 and 7.7 +/- 0.1 for cleavage and 6.1 +/- 0.3 and 6.9 +/- 0.3 for ligation. The data provide good evidence for general acid-base catalysis by the nucleobases.
Figures
References
-
- Araki L., Harusawa S., Yamaguchi M., Yonezawa S., Taniguchi N., Lilley D.M.J., Zhao Z., Kurihara T. Synthesis of C4-linked imidazole ribonucleoside phosphoramidite with pivaloyloxymethyl (POM) group. Tetrahedron Lett. 2004;45:2657–2661.
-
- Bevilacqua P.C. Mechanistic considerations for general acid-base catalysis by RNA: Revisiting the mechanism of the hairpin ribozyme. Biochemistry. 2003;42:2259–2265. - PubMed
-
- Bevilacqua P.C. Mechanism of catalytic RNA. Biopolymers. 2004;73:69–70. - PubMed
-
- Clegg R.M. Fluorescence resonance energy transfer and nucleic acids. Methods Enzymol. 1992;211:353–388. - PubMed
-
- Fedor M.J. Tertiary structure stabilization promotes hairpin ribozyme ligation. Biochemistry. 1999;38:11040–11050. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources